
Contents
- 1 教科書
- 1.1 Objectives
- 1.2 生物医学的重要性 (Medical Importance)
- 1.3 脂肪酸の新規合成経路(Main Pathway for De Novo Synthesis of Fatty Acids, Lipogenesis)は細胞質(cytosol)で行われる
- 1.4 マロニルCoA(Malonyl-CoA)の生成は脂肪酸合成の最初かつ制御段階である
- 1.5 脂肪酸合成酵素複合体(Fatty Acid Synthase Complex)は6つの酵素活性とアシルキャリアタンパク質(Acyl Carrier Protein, ACP)を含む2つのポリペプチド鎖からなるホモダイマー(homodimer)である
- 1.6 マロニルCoA(Malonyl-CoA)の形成後の脂肪酸の合成
- 1.7 脂肪酸合成の反応機構
- 1.8 パルミチン酸の活性化とその運命
- 1.9 パルミチン酸の合成全体の化学反応式
- 1.10 奇数炭素鎖脂肪酸の合成
- 1.11 脂肪生成(Lipogenesis)におけるNADPHの主な供給源はペントースリン酸経路(Pentose Phosphate Pathway)である
- 1.12 アセチルCoA(Acetyl-CoA)は脂肪酸の主要構成要素である
- 1.13 クエン酸輸送とマレートの役割
- 1.14 反芻動物(Ruminants)の例外
- 1.15 脂肪酸鎖の伸長は小胞体で行われる
- 1.16 栄養状態が脂肪生成(Lipogenesis)を調節する
- 1.17 脂肪生成の短期および長期の調節機構
- 1.18 アセチルCoAカルボキシラーゼ(Acetyl-CoA Carboxylase)は脂肪生成の調節で最も重要な酵素である
- 1.19 ホルモンによる調節
- 1.20 ピルビン酸デヒドロゲナーゼ(Pyruvate Dehydrogenase)はアシルCoA(Acyl-CoA)によっても調節される
- 1.21 インスリン(Insulin)はその他のメカニズムによっても脂肪生成を調節する
- 1.22 脂肪酸合成酵素複合体(Fatty Acid Synthase Complex)およびアセチルCoAカルボキシラーゼ(Acetyl-CoA Carboxylase)は適応酵素である
- 1.23 多価不飽和脂肪酸(Polyunsaturated Fatty Acids)は哺乳類では合成できず、栄養的に必須である
- 1.24 一価不飽和脂肪酸(Monounsaturated Fatty Acids)はΔ9デサチュラーゼ系(Δ9 Desaturase System)によって合成される
- 1.25 多価不飽和脂肪酸(Polyunsaturated Fatty Acids)の合成にはデサチュラーゼ(Desaturase)およびエロンゲース(Elongase)酵素系が関与する
- 1.26 必須脂肪酸(Essential Fatty Acids, EFA)は体内で重要な役割を果たす
- 1.27 エイコサノイド(Eicosanoids)はC20多価不飽和脂肪酸から形成される
- 1.28 シクロオキシゲナーゼ経路(Cyclooxygenase Pathway)はプロスタノイド(Prostanoids)の合成を担う
- 1.29 ロイコトリエン(Leukotrienes)およびリポキシン(Lipoxins)はリポキシゲナーゼ経路(Lipoxygenase Pathway)で形成される
- 1.30 臨床的側面(Clinical Aspects)
- 1.31 必須脂肪酸欠乏症(Essential Fatty Acid Deficiency)の症状には皮膚病変(Skin Lesions)と脂質輸送の障害(Impairment of Lipid Transport)が含まれる
- 1.32 プロスタノイド(Prostanoids)の治療用途
- 1.33 ロイコトリエン(Leukotrienes)とリポキシン(Lipoxins)は多くの疾患プロセスの調節因子
- 1.34 必須脂肪酸(Essential Fatty Acids)の異常代謝
- 1.35 トランス脂肪酸(Trans Fatty Acids)はさまざまな障害に関連
- 1.36 非ステロイド性抗炎症薬(NSAIDs)はCOXを阻害する
- 2 重要箇所穴埋め問題
教科書

Objectives
1. アセチルCoAカルボキシラーゼ(Acetyl-CoA Carboxylase)が触媒する反応とその調節機構
- 反応の内容:
- アセチルCoA(acetyl-CoA)をカルボキシル化し、マロニルCoA(malonyl-CoA)を生成します。この反応は脂肪酸合成の律速段階(rate-limiting step)です。
- 反応に必要な要素:
- ATP
- ビオチン(biotin, ビタミンB7)
- 2段階のプロセス:
- ビオチンがATPを利用してカルボキシル化。
- カルボキシル基がアセチルCoAに転移され、マロニルCoAを生成。
- 調節機構:
- アロステリック調節(Allosteric Regulation):
- クエン酸(citrate)は酵素を活性化。
- 長鎖アシルCoA(long-chain acyl-CoA)は酵素を阻害。
- ホルモン調節(Hormonal Regulation):
- インスリン(insulin)は酵素を脱リン酸化し、活性化。
- グルカゴン(glucagon)とエピネフリン(epinephrine)はリン酸化を促進し、活性を低下。
- アロステリック調節(Allosteric Regulation):
2. 脂肪酸合成酵素複合体(Fatty Acid Synthase Complex)の構造
- 構造:
- ホモダイマー(homodimer)で、各ペプチド鎖に6つの酵素活性部位とアシルキャリアタンパク質(acyl carrier protein, ACP)が含まれる。
- 主な酵素の順序:
- アセチル/マロニルトランスアシラーゼ(Acetyl/Malonyl Transacylase)
- 3-ケトアシル合成酵素(3-Ketoacyl Synthase)
- 3-ケトアシル還元酵素(3-Ketoacyl Reductase)
- デヒドラターゼ(Dehydratase)
- エノイル還元酵素(Enoyl Reductase)
- チオエステラーゼ(Thioesterase)
- 特徴:
- ACPは脂肪酸の伸長に必要なパントテン酸(pantothenic acid)の誘導体を含む。
- 全体的な構造はX型で、効率的な酵素活性を可能にする。
3. 長鎖脂肪酸の合成と16炭素パルミチン酸の形成
- メカニズム:
- マロニルCoAからの2炭素単位の反復縮合(condensation)によって脂肪酸鎖が伸長。
- 16炭素のパルミチン酸(palmitate)の合成が最も一般的。
- 必要な補因子:
- NADPH(還元等価物、reducing equivalents)
- ATP
- ビオチン(biotin)
4. 脂肪酸合成に必要な還元等価物(NADPH)の供給源
- 主な供給源:
- ペントースリン酸経路(Pentose Phosphate Pathway):
- グルコース6-リン酸デヒドロゲナーゼ(glucose-6-phosphate dehydrogenase)の活性によりNADPHを生成。
- リンゴ酸シャトル(Malic Enzyme Pathway):
- リンゴ酸(malate)の酸化的脱炭酸によりNADPHを生成。
- ペントースリン酸経路(Pentose Phosphate Pathway):
5. 脂肪酸合成の栄養状態による調節
- 栄養状態の影響:
- 高炭水化物食(high-carbohydrate diet):
- インスリンによるアセチルCoAカルボキシラーゼと脂肪酸合成酵素複合体の活性化。
- 飢餓状態(fasting):
- グルカゴンとエピネフリンによる阻害。
- 高炭水化物食(high-carbohydrate diet):
- 追加の制御機構:
- 遺伝子発現(gene expression)の調節。
- マロニルCoAがカルニチンパルミトイルトランスフェラーゼ1(CPT1)を阻害し、脂肪酸酸化を制限。
6. 栄養的に必須な脂肪酸
- 必須脂肪酸(Essential Fatty Acids):
- リノール酸(linoleic acid, ω-6脂肪酸)
- α-リノレン酸(α-linolenic acid, ω-3脂肪酸)
- 体内で合成できない理由:
- ヒトはΔ12およびΔ15不飽和酵素(desaturase enzymes)を持たないため、これらの多価不飽和脂肪酸(PUFAs)を食事から摂取する必要がある。
7. 多価不飽和脂肪酸の合成
- 酵素の関与:
- デサチュラーゼ(Desaturase):
- 脂肪酸に二重結合を導入。
- エロンゲーション酵素(Elongation Enzymes):
- 脂肪酸鎖を伸長。
- デサチュラーゼ(Desaturase):
- 主なステップ:
- Δ9デサチュラーゼによる二重結合の挿入(例:ステアリン酸→オレイン酸)。
8. エイコサノイド(Eicosanoids)の合成経路
- シクロオキシゲナーゼ経路(Cyclooxygenase Pathway):
- プロスタグランジン(prostaglandins)とトロンボキサン(thromboxanes)を生成。
- 炎症、痛み、血小板凝集の調節に関与。
- NSAIDs(例:アスピリン)はこの経路を阻害。
- リポキシゲナーゼ経路(Lipoxygenase Pathway):
- ロイコトリエン(leukotrienes)とリポキシン(lipoxins)を生成。
- アレルギー反応や炎症に重要。
生物医学的重要性 (Medical Importance)
脂肪酸(fatty acids)は、細胞質(cytosol)におけるミトコンドリア外(extramitochondrial)システムによって合成されます。このシステムは、アセチルCoA(acetyl-CoA)からパルミチン酸(palmitate)を完全に合成する役割を果たします。ほとんどの哺乳類では、グルコース(glucose)が脂肪生成(lipogenesis)の主要基質ですが、反芻動物(ruminants)では酢酸(acetate)が主要燃料分子です。これらは食事から得られます。この経路に関連する重大な疾患はヒトでは報告されていませんが、1型糖尿病(type 1 diabetes mellitus, insulin-dependent diabetes)では脂肪生成の抑制が起こります。また、このプロセスの活性の変動は肥満(obesity)の性質や程度に影響を与えます。
細胞膜(cell membrane)のリン脂質(phospholipids)中の不飽和脂肪酸(unsaturated fatty acids)は、膜流動性(membrane fluidity)の維持に重要です(第40章参照)。食事中の多価不飽和脂肪酸(polyunsaturated fatty acids, PUFAs)と飽和脂肪酸(saturated fatty acids, SFAs)の比率(P:S比)が高いことは、冠動脈性心疾患(coronary heart disease)の予防に有益とされています。動物組織は脂肪酸の不飽和化能力が限られているため、植物由来の多価不飽和脂肪酸が必要です。これらの必須脂肪酸(essential fatty acids)は、エイコサノイド(eicosanoids)の前駆体となるエイコサン酸(C20脂肪酸)を形成します。このエイコサノイドには、プロスタグランジン(prostaglandins)、トロンボキサン(thromboxanes)、ロイコトリエン(leukotrienes)、およびリポキシン(lipoxins)が含まれます。
プロスタグランジンは炎症(inflammation)、痛み(pain)、睡眠(sleep)を媒介し、血液凝固(blood coagulation)や生殖(reproduction)を調節します。一方、非ステロイド性抗炎症薬(NSAIDs, nonsteroidal anti-inflammatory drugs)であるアスピリン(aspirin)やイブプロフェン(ibuprofen)は、プロスタグランジン合成を阻害することで作用します。ロイコトリエンは筋収縮(muscle contractant)や化学走性(chemotaxis)を持ち、アレルギー反応(allergic reactions)や炎症において重要です。
脂肪酸の新規合成経路(Main Pathway for De Novo Synthesis of Fatty Acids, Lipogenesis)は細胞質(cytosol)で行われる
このシステムは、肝臓(liver)、腎臓(kidney)、脳(brain)、肺(lung)、乳腺(mammary gland)、および脂肪組織(adipose tissue)を含む多くの組織に存在します。その補因子(cofactor)には、NADPH、ATP、Mn²⁺、ビオチン(biotin)、およびHCO₃⁻(CO₂の供給源として)が含まれます。アセチルCoAが直接的な基質であり、遊離パルミチン酸(free palmitate)が最終生成物です。
マロニルCoA(Malonyl-CoA)の生成は脂肪酸合成の最初かつ制御段階である
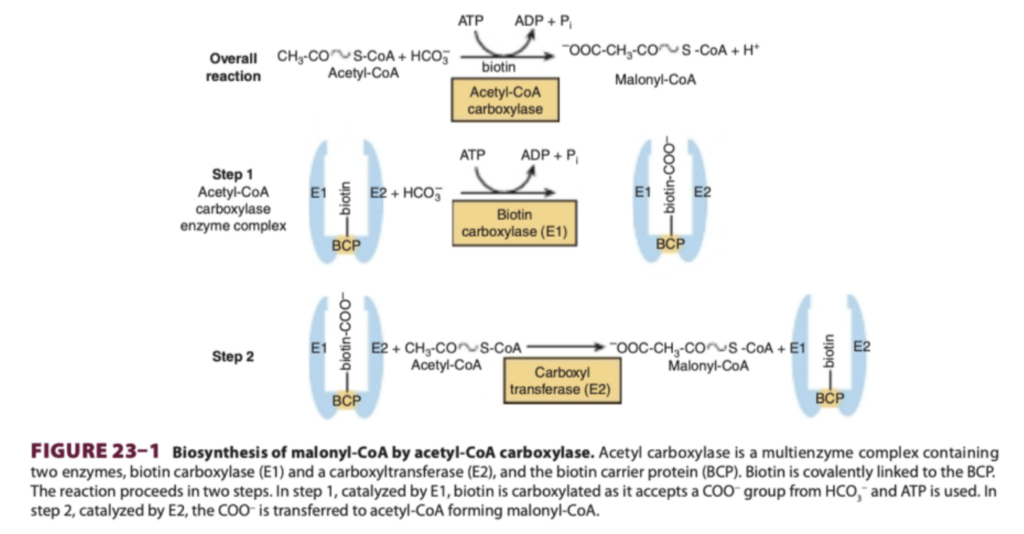
脂肪酸合成の最初のステップは、アセチルCoA(acetyl-CoA)のカルボキシル化(carboxylation)によりマロニルCoA(malonyl-CoA)を形成することです。この反応はアセチルCoAカルボキシラーゼ(acetyl-CoA carboxylase)によって触媒されます。この反応ではATPとビタミンBの一種であるビオチン(biotin)が必要です。
アセチルCoAカルボキシラーゼは以下を含む多酵素タンパク質です:
- ビオチンカルボキシラーゼ(biotin carboxylase)
- カルボキシル基転移酵素(carboxyl transferase)
- ビオチン結合タンパク質(biotin carrier protein, ビオチンを結合する)
また、調節のためのアロステリック部位(allosteric site)も含まれています。この反応は以下の2段階で進行します:
- 第1段階: ビオチンカルボキシラーゼが触媒し、ビオチンをカルボキシル化。ATPを使用します。
- 第2段階: カルボキシル基転移酵素が触媒し、カルボキシル基をアセチルCoAに転移してマロニルCoAを形成します。
アセチルCoAカルボキシラーゼは、脂肪酸合成の調節において主要な役割を果たします(詳細は後述します)。
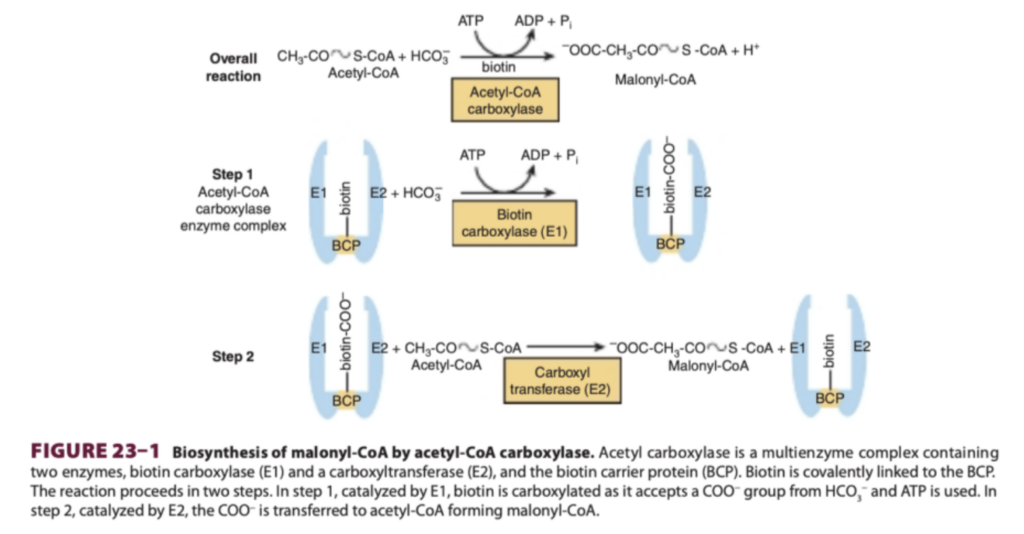
脂肪酸合成酵素複合体(Fatty Acid Synthase Complex)は6つの酵素活性とアシルキャリアタンパク質(Acyl Carrier Protein, ACP)を含む2つのポリペプチド鎖からなるホモダイマー(homodimer)である
マロニルCoA(Malonyl-CoA)の形成後の脂肪酸の合成
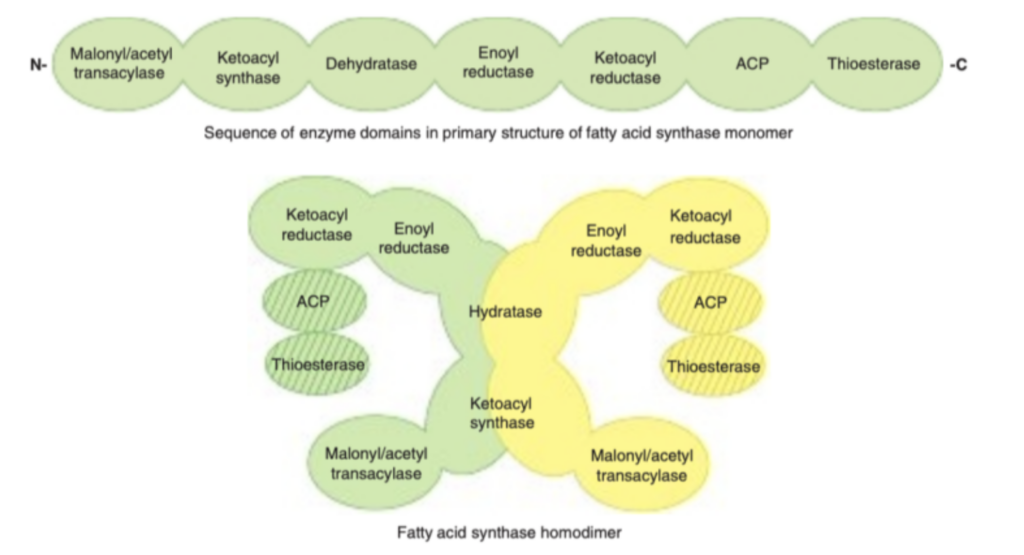
- 概要: 脂肪酸は脂肪酸合成酵素複合体(fatty acid synthase enzyme complex)によって形成されます。この複合体には、脂肪酸合成に必要な各酵素がリンクされた状態で存在し、アシルキャリアタンパク質(acyl carrier protein, ACP)も組み込まれています。ACPは、β酸化経路(β-oxidation pathway, 第22章参照)におけるCoAと同様の機能を果たします。
- 重要な構造的特徴:
- ACPは、4′-ホスホパンテテイン(4′-phosphopantetheine)の形でビタミンパントテン酸(pantothenic acid)を含む。
- タンパク質の一次構造では酵素ドメインが順番に結合しているとされますが、X線結晶解析(X-ray crystallography)により、この複合体が「X型」のホモダイマー構造を持つことが示されています。
- 各サブユニットには6つの酵素活性部位と1つのACPが含まれており、細胞内で透過性障壁を必要としない効率的なプロセスの区画化を可能にしています。また、これらすべての酵素は単一の遺伝子でコードされています。
脂肪酸合成の反応機構
- 初期段階:
- アセチルCoA(acetyl-CoA)のプライミング分子が、複合体の1つのモノマー内にあるシステイン(cysteine)の—SH基と結合。
- 一方、マロニルCoA(malonyl-CoA)は、隣接するモノマーのACP上の4′-ホスホパンテテインの—SH基と結合。
- 触媒反応:
- マロニルアセチルトランスアシラーゼ(Malonyl Acetyl Transacylase):
- アセチル(アシル)-マロニル酵素を形成。
- 3-ケトアシル合成酵素(3-Ketoacyl Synthase):
- アセチル基がマロニル基のメチレン基を攻撃し、CO₂を放出して3-ケトアシル酵素(acetoacetyl enzyme)を形成。
- 脱炭酸反応(Decarboxylation):
- この反応によりエネルギーが放出され、全ての反応が前方に進行。
- マロニルアセチルトランスアシラーゼ(Malonyl Acetyl Transacylase):
- 還元と脱水:
- 3-ケトアシル還元酵素(3-Ketoacyl Reductase):
- 還元反応を触媒。
- デヒドラターゼ(Dehydratase):
- 脱水反応を触媒。
- エノイル還元酵素(Enoyl Reductase):
- 再度還元反応を触媒し、飽和アシル酵素を形成。
- 3-ケトアシル還元酵素(3-Ketoacyl Reductase):
- 鎖伸長の繰り返し:
- 飽和アシル基はACPの—SH基からシステインの—SH基へ転移し、新しいマロニルCoAが結合します。
- この反応は6回繰り返され、16炭素のパルミトイル(palmitoyl)が組み立てられます。
- 最終段階:
- チオエステラーゼ(Thioesterase, Deacylase):
- パルミチン酸(palmitate)を複合体から遊離させます。
- チオエステラーゼ(Thioesterase, Deacylase):
パルミチン酸の活性化とその運命
- 活性化: 遊離パルミチン酸はアシルCoA(acyl-CoA)として活性化されます。
- 可能な運命:
- アシルグリセロール(acylglycerols)へのエステル化(esterification)
- 鎖の伸長(chain elongation)
- 不飽和化(desaturation)
- コレステリルエステル(cholesteryl ester)へのエステル化
- 乳腺(mammary gland)での特例:
- 乳腺にはC8、C10、C12のアシル残基に特異的なチオエステラーゼが存在し、これらは乳脂肪中に含まれます。
パルミチン酸の合成全体の化学反応式
CH3CO-S-CoA+7HOOCCHCO-S-CoA+14NADPH+14H+
→CH3(CH2)14COOH+7CO2+6H2O+8CoA-SH+14NADP
- 詳細:
- アセチルCoAはパルミチン酸の炭素15と16を形成。
- それ以外の炭素はすべてマロニルCoA由来。
奇数炭素鎖脂肪酸の合成
- 特徴: アセチルCoAの代わりにプロピオニルCoA(propionyl-CoA)がプライマーとして使用されます。
- 例: 奇数炭素鎖脂肪酸は特に反芻動物(ruminants)の脂肪や乳中に見られます。
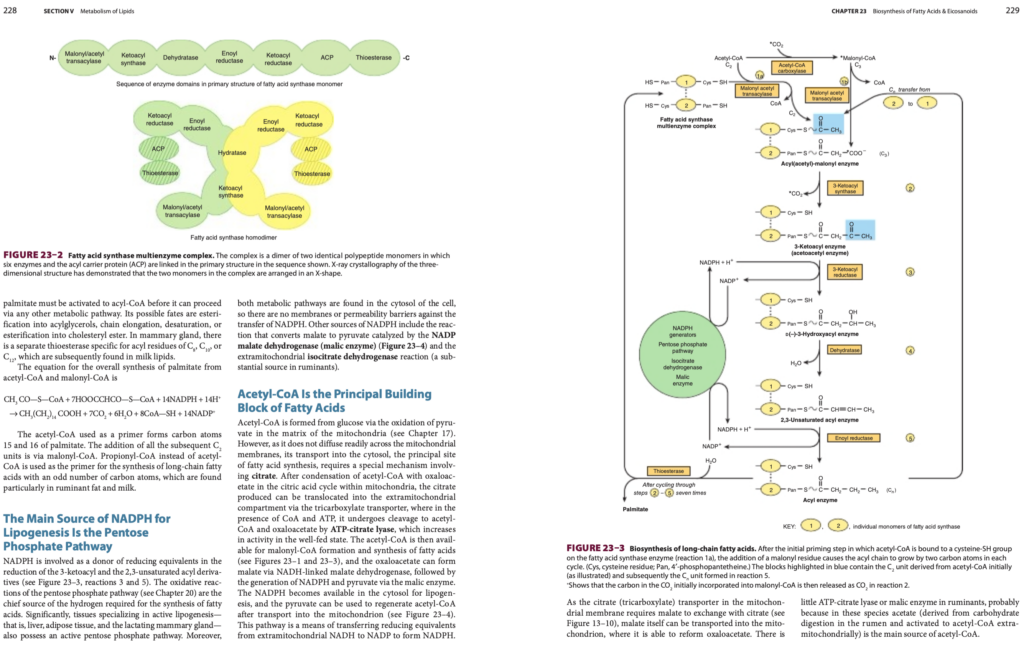
脂肪生成(Lipogenesis)におけるNADPHの主な供給源はペントースリン酸経路(Pentose Phosphate Pathway)である
NADPHは、3-ケトアシル(3-ketoacyl)および2,3-不飽和アシル誘導体(2,3-unsaturated acyl derivatives)の還元反応(reduction reactions)において、還元当量(reducing equivalents)の供与体として関与します(図23–3、反応3および5参照)。ペントースリン酸経路(pentose phosphate pathway, 第20章参照)の酸化反応(oxidative reactions)は、脂肪酸合成(fatty acid synthesis)に必要な水素(hydrogen)の主な供給源です。
特に、活発な脂肪生成を行う組織(肝臓[liver]、脂肪組織[adipose tissue]、授乳中の乳腺[lactating mammary gland])は、活性化されたペントースリン酸経路を持っています。さらに、これらの代謝経路は細胞質(cytosol)内に存在するため、NADPHの移動を妨げる膜や透過性障壁が存在しません。
NADPHの他の供給源:
- マレート(malate)をピルビン酸(pyruvate)に変換する反応(NADP-マレートデヒドロゲナーゼ[NADP malate dehydrogenase, マレック酵素(malic enzyme)]による触媒)。
- ミトコンドリア外のイソクエン酸デヒドロゲナーゼ(extramitochondrial isocitrate dehydrogenase)反応(特に反芻動物[ruminants]での重要な供給源)。
アセチルCoA(Acetyl-CoA)は脂肪酸の主要構成要素である
アセチルCoAは、ピルビン酸(pyruvate)の酸化(oxidation)を通じてグルコース(glucose)から生成され、ミトコンドリア(mitochondria)のマトリックス(matrix)内で形成されます(第17章参照)。しかし、アセチルCoAはミトコンドリア膜(mitochondrial membrane)を容易には拡散しないため、脂肪酸合成の主要な場である細胞質(cytosol)への輸送には特別な機構が必要です。この機構にはクエン酸(citrate)が関与します。
- クエン酸を介した輸送機構:
- ミトコンドリア内でアセチルCoAがオキサロ酢酸(oxaloacetate)と縮合し、クエン酸回路(citric acid cycle)でクエン酸を生成。
- 生成されたクエン酸は三カルボン酸トランスポーター(tricarboxylate transporter)によってミトコンドリア外に移動。
- 細胞質では、クエン酸がCoAおよびATPの存在下でATP-クエン酸リアーゼ(ATP-citrate lyase)によってアセチルCoAとオキサロ酢酸に分解される。この酵素の活性は栄養が十分な状態(well-fed state)で増加。
- その後の代謝プロセス:
- アセチルCoAは、マロニルCoA(malonyl-CoA)の形成および脂肪酸合成に利用。
- オキサロ酢酸はNADHに依存したマレートデヒドロゲナーゼ(malate dehydrogenase)によってマレートを形成。
- マレートはマレック酵素(malic enzyme)を介してNADPHおよびピルビン酸を生成。
- 生成されたNADPHは細胞質で脂肪生成に利用され、ピルビン酸はミトコンドリア内に輸送され、再びアセチルCoAを再生。
この経路は、ミトコンドリア外NADHからNADPへの還元当量(reducing equivalents)の移動を通じてNADPHを生成する手段を提供します。
クエン酸輸送とマレートの役割
- 三カルボン酸トランスポーター(tricarboxylate transporter)は、クエン酸の輸送のためにマレートを必要とします(図13–10参照)。
- マレートはミトコンドリア内に輸送され、オキサロ酢酸に再変換されることが可能。
反芻動物(Ruminants)の例外
- 反芻動物ではATP-クエン酸リアーゼ(ATP-citrate lyase)やマレック酵素(malic enzyme)の活性が低い。
- 主な理由は、これらの動物では脂肪酸合成の主要なアセチルCoAの供給源が酢酸(acetate)であること。酢酸はルーメン(rumen)での炭水化物消化から得られ、ミトコンドリア外でアセチルCoAに活性化されます。
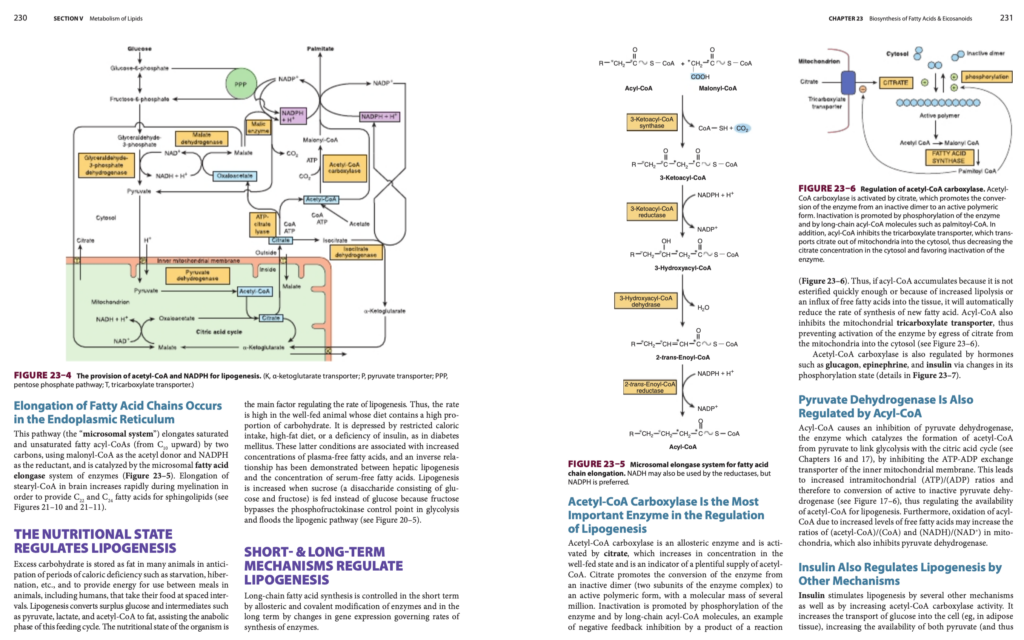
脂肪酸鎖の伸長は小胞体で行われる
この経路(「ミクロソーム系」)は、飽和および不飽和アシル-CoA(C10以上)を2炭素分伸長させるもので、マロニルCoA(malonyl-CoA)をアセチル供与体として使用し、NADPHを還元剤として利用します。この反応は、ミクロソーム脂肪酸伸長酵素系(microsomal fatty acid elongase system)によって触媒されます(図23–5参照)。脳内でのステアリル-CoA(stearyl-CoA)の伸長は、髄鞘形成(myelination)中に急速に増加し、スフィンゴ脂質(sphingolipids)に必要なC22およびC24の脂肪酸を供給します(図21–10および21–11参照)。
栄養状態が脂肪生成(Lipogenesis)を調節する
過剰な炭水化物は多くの動物で脂肪として蓄えられ、飢餓(starvation)や冬眠(hibernation)などのカロリー不足の期間や、食事が間隔を空けて摂取される動物(ヒトを含む)での食事と食事の間のエネルギー供給に備えます。脂肪生成は、余分なグルコース(glucose)やピルビン酸(pyruvate)、乳酸(lactate)、アセチルCoA(acetyl-CoA)などの中間産物を脂肪に変換し、この摂食サイクルの同化的(anabolic)な段階を助けます。
脂肪生成の速度は、主に生物の栄養状態によって調節されます。栄養状態が良好な動物、特に炭水化物が多い食事を取っている場合、脂肪生成は高くなります。制限されたカロリー摂取、高脂肪食、またはインスリンの欠乏(糖尿病など)によって脂肪生成は抑制されます。これらの後者の状態では、血漿中の遊離脂肪酸(free fatty acids)の濃度が増加し、肝臓での脂肪生成と血清中の遊離脂肪酸濃度との間に逆相関(inverse relationship)が示されています。
また、グルコースの代わりにショ糖(sucrose、グルコースとフルクトースからなる二糖類)を与えると脂肪生成が増加します。フルクトースは解糖系(glycolysis)のホスホフルクトキナーゼ制御点(phosphofructokinase control point)を回避し、脂肪生成経路を飽和させます(図20–5参照)。
脂肪生成の短期および長期の調節機構
長鎖脂肪酸の合成は、酵素のアロステリックおよび共役的修飾(covalent modification)によって短期的に制御され、遺伝子発現の変化によって長期的に制御されます。これらは酵素の合成速度に影響を与えます。
アセチルCoAカルボキシラーゼ(Acetyl-CoA Carboxylase)は脂肪生成の調節で最も重要な酵素である
アセチルCoAカルボキシラーゼはアロステリック酵素であり、クエン酸(citrate)によって活性化されます。クエン酸は、栄養が十分な状態(well-fed state)でその濃度が増加し、アセチルCoAの供給が豊富であることを示す指標となります。クエン酸は、酵素を不活性なダイマー(酵素複合体の2つのサブユニット)から、数百万の分子量を持つ活性化された重合体(polymeric form)に変換するのを促進します。
不活性化のメカニズム:
- 酵素のリン酸化(phosphorylation)や、長鎖アシル-CoA(long-chain acyl-CoA)分子によって不活性化が促進されます。これは、反応産物による負のフィードバック抑制(negative feedback inhibition)の一例です(図23–6参照)。
- したがって、アシル-CoAが蓄積し、十分にエステル化されない場合や、脂肪分解の増加、遊離脂肪酸の組織への流入が起こると、脂肪酸の新たな合成速度が自動的に減少します。
アシル-CoAの役割:
- アシル-CoAはミトコンドリア内の三カルボン酸トランスポーター(tricarboxylate transporter)も抑制し、クエン酸がミトコンドリアから細胞質へ移行するのを防ぎます(図23–6参照)。
ホルモンによる調節
アセチルCoAカルボキシラーゼは、グルカゴン(glucagon)、エピネフリン(epinephrine)、インスリン(insulin)などのホルモンによっても調節されます。これらのホルモンは、酵素のリン酸化状態に変化をもたらします(図23–7参照)。
ピルビン酸デヒドロゲナーゼ(Pyruvate Dehydrogenase)はアシルCoA(Acyl-CoA)によっても調節される
アシルCoAはピルビン酸デヒドロゲナーゼ(pyruvate dehydrogenase)を抑制します。この酵素は、解糖系(glycolysis)とクエン酸回路(citric acid cycle)を結びつける役割を果たすピルビン酸からアセチルCoA(acetyl-CoA)への変換を触媒します(第16章および第17章参照)。アシルCoAは、ミトコンドリア内膜(inner mitochondrial membrane)のATP-ADP交換トランスポーター(ATP-ADP exchange transporter)を阻害することでこの抑制を引き起こします。
これにより、ミトコンドリア内のATP/ADP比((ATP)/(ADP) ratio)が増加し、活性型ピルビン酸デヒドロゲナーゼ(active pyruvate dehydrogenase)が不活性型(inactive form)に変換されます(図17–6参照)。このメカニズムによって、脂肪生成(lipogenesis)に利用可能なアセチルCoAの供給が調節されます。
さらに、遊離脂肪酸(free fatty acids)の濃度が増加することでアシルCoAの酸化(oxidation of acyl-CoA)が進行すると、ミトコンドリア内のアセチルCoA/CoA比((acetyl-CoA)/(CoA) ratio)およびNADH/NAD⁺比((NADH)/(NAD⁺) ratio)が増加し、これもピルビン酸デヒドロゲナーゼを抑制します。
インスリン(Insulin)はその他のメカニズムによっても脂肪生成を調節する
インスリンはアセチルCoAカルボキシラーゼ(acetyl-CoA carboxylase)の活性を増加させるだけでなく、その他のメカニズムによって脂肪生成を刺激します。
- 細胞へのグルコース輸送の増加:
- 例:脂肪組織(adipose tissue)においてグルコースの細胞内輸送を促進。
- ピルビン酸(pyruvate)およびそれに続くアセチルCoA(acetyl-CoA)の供給が増加し、脂肪酸合成(fatty acid synthesis)が促進されます。
- また、新たに形成された脂肪酸のエステル化(esterification)を介してトリアシルグリセロール(triacylglycerol)合成のためのグリセロール-3-リン酸(glycerol-3-phosphate)も増加します(図24–2参照)。
- ピルビン酸デヒドロゲナーゼの活性化:
- インスリンは、脂肪組織において不活性型ピルビン酸デヒドロゲナーゼを活性型に変換します(肝臓ではこの作用は見られません)。
- リポリシス(脂肪分解)の抑制:
- インスリンは細胞内cAMP濃度(intracellular cAMP levels)を低下させる能力を持ち、脂肪組織での脂肪分解を阻害。
- これにより、血漿遊離脂肪酸(plasma-free fatty acids)の濃度および長鎖アシルCoA(long-chain acyl-CoA)の濃度が減少します。
- 長鎖アシルCoAは脂肪生成の阻害因子であるため、この濃度低下により脂肪生成が促進されます。

脂肪酸合成酵素複合体(Fatty Acid Synthase Complex)およびアセチルCoAカルボキシラーゼ(Acetyl-CoA Carboxylase)は適応酵素である
これらの酵素は、遺伝子発現(gene expression)の変化を通じて、体の生理的な必要性に適応します。これにより、栄養が十分な状態(fed state)では酵素タンパク質の総量が増加し、高脂肪食、飢餓(starvation)、および糖尿病(diabetes mellitus)などの条件では減少します。
- インスリン(Insulin)の役割:
- インスリンは、遺伝子発現と酵素生合成(enzyme biosynthesis)の誘導を促進します。
- グルカゴン(Glucagon)の役割:
- cAMPを介してインスリンの作用を拮抗します。
- 多価不飽和脂肪酸(polyunsaturated fatty acids)を含む脂肪の摂取:
- 解糖系(glycolysis)と脂肪生成経路(lipogenesis)の主要酵素の発現を抑制する調節機構を持つ。
これらの脂肪生成の長期的な調節メカニズムが完全に発現するには数日かかります。また、遊離脂肪酸(free fatty acids)やインスリン、グルカゴンなどのホルモンの即時かつ直接的な作用を補完します。
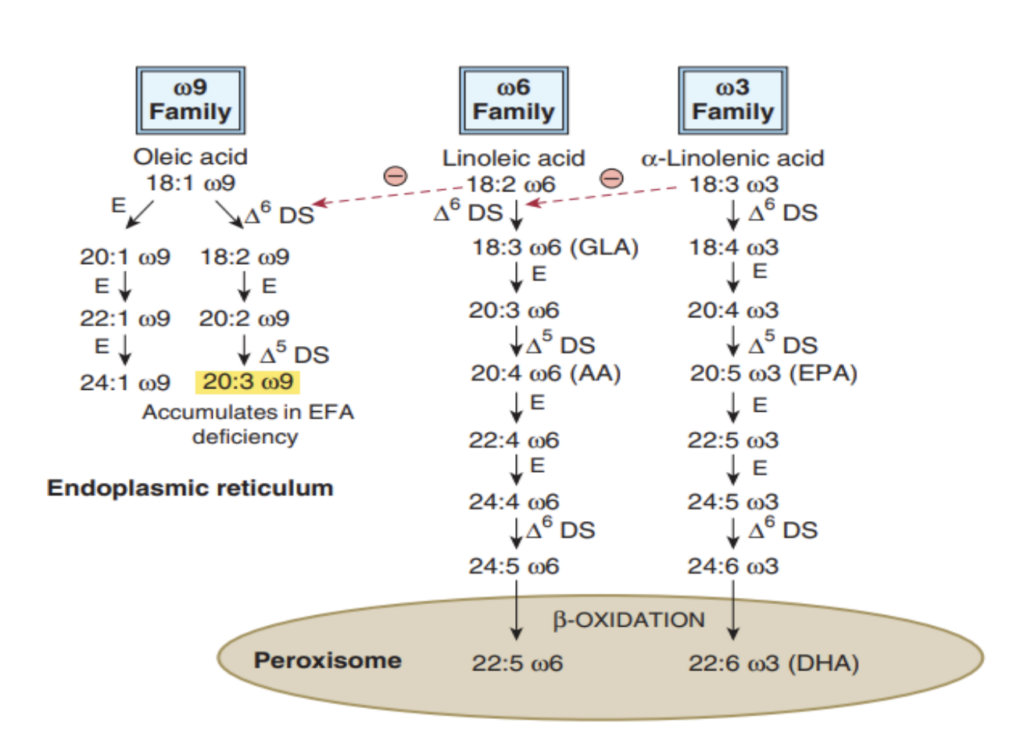
多価不飽和脂肪酸(Polyunsaturated Fatty Acids)は哺乳類では合成できず、栄養的に必須である
- 哺乳類における代謝的に重要な特定の長鎖不飽和脂肪酸(long-chain unsaturated fatty acids)は、図23–8に示されています。
- オレイン酸(oleic acid)、
リノール酸(linoleic acid)、
α-リノレン酸(α-linolenic acid)
から、鎖伸長(chain elongation)を通じて他のC20、C22、C24の多価脂肪酸が生成されます。
必須脂肪酸(Essential Fatty Acids):
- リノール酸(linoleic acid)およびα-リノレン酸(α-linolenic acid)は、多くの動物種(ヒトを含む)における完全な栄養に必須であり、栄養的必須脂肪酸(nutritionally essential fatty acids)と呼ばれます。
- ヒトやほとんどの哺乳類では、リノール酸からアラキドン酸(arachidonic acid)が形成されます。
二重結合の導入:
- 多くの動物では、Δ4、Δ5、Δ6、およびΔ9位置に二重結合を導入可能です(第21章参照)。ただし、Δ9を超える位置には導入できません。
- 植物はΔ12およびΔ15位置に二重結合を導入できるため、必須脂肪酸を合成できます。
一価不飽和脂肪酸(Monounsaturated Fatty Acids)はΔ9デサチュラーゼ系(Δ9 Desaturase System)によって合成される
- 飽和脂肪酸(saturated fatty acids)から一価不飽和脂肪酸(monounsaturated fatty acids)を生成する主な組織には肝臓(liver)などがあります。
- 飽和脂肪酸に最初に導入される二重結合は、ほぼ常にΔ9位置です。
Δ9デサチュラーゼ系(Δ9 Desaturase System):
- 小胞体(endoplasmic reticulum)内の酵素系が、パルミトイルCoA(palmitoyl-CoA)やステアロイルCoA(stearoyl-CoA)をそれぞれパルミトレオイルCoA(palmitoleoyl-CoA)やオレオイルCoA(oleoyl-CoA)に変換します(図23–9参照)。
パルミトイルCoA(palmitoyl-CoA)
→パルミトレオイルCoA(palmitoleoyl-CoA)
ステアロイルCoA(stearoyl-CoA)
→オレオイルCoA(oleoyl-CoA) - 酸素(oxygen)とNADHまたはNADPHが反応に必要です。
- 酵素は、シトクロムb5(cytochrome b5)を含むモノオキシゲナーゼ系(mono-oxygenase system, 第12章参照)に類似しています。
多価不飽和脂肪酸(Polyunsaturated Fatty Acids)の合成にはデサチュラーゼ(Desaturase)およびエロンゲース(Elongase)酵素系が関与する
- 既存の一価不飽和脂肪酸にさらに二重結合を導入することで、多価不飽和脂肪酸が形成されます。
- 特徴:
- 動物では、二重結合はメチレン基(methylene group)で分離されています(細菌では例外)。
- Δ9デサチュラーゼを持つ動物は、鎖伸長(chain elongation)と不飽和化(desaturation)の組み合わせを通じて、ω9(オレイン酸)脂肪酸ファミリーを完全に合成可能です(図23–9および23–10参照)。
必須脂肪酸の重要性:
- リノール酸(ω6)およびα-リノレン酸(ω3)は、これらのファミリーの他のメンバーを合成するために必要であり、食事から摂取する必要があります。
- 例:
- リノール酸はγ-リノレン酸(18:3 ω6)を経てアラキドン酸(20:4 ω6)に変換されます。
例外:
- 猫(cats)はΔ6デサチュラーゼ(Δ6 desaturase)を持たないため、アラキドン酸を食事から摂取する必要があります。
調節:
- デサチュレーションと鎖伸長システムは、飢餓状態(starvation)、グルカゴン(glucagon)やエピネフリン(epinephrine)の投与、インスリン(insulin)の欠如(1型糖尿病)により大幅に減少します。

必須脂肪酸(Essential Fatty Acids, EFA)は体内で重要な役割を果たす
- 動物実験の結果:
- ビタミンAおよびDを含む純粋な非脂質食(nonlipid diet)を与えられたラットでは、成長速度の低下と生殖障害が観察されますが、食事にリノール酸(linoleic acid)、α-リノレン酸(α-linolenic acid)、およびアラキドン酸(arachidonic acid)を加えることで改善されます。
- これらの脂肪酸は植物油(vegetable oils, 表21–2参照)に高濃度で含まれ、動物性脂肪には少量含まれています。
- 役割:
- プロスタグランジン(prostaglandins)、
トロンボキサン(thromboxanes)、
ロイコトリエン(leukotrienes)、
リポキシン(lipoxins)
の形成に必要。 - 細胞の構造脂質(structural lipids)に含まれ、多くの場合リン脂質(phospholipids)の位置2に存在。
- ミトコンドリア膜(mitochondrial membrane)の構造的完全性を維持。
- プロスタグランジン(prostaglandins)、
- アラキドン酸(arachidonic acid):
- 細胞膜に存在し、リン脂質中の脂肪酸の5–15%を占める。
- ドコサヘキサエン酸(DHA; ω3, 22:6):
- α-リノレン酸からわずかに合成されるか、魚油から直接得られる。
- 網膜(retina)、大脳皮質(cerebral cortex)、精巣(testis)、精子(sperm)に高濃度で存在し、脳と網膜の発達に必要。
- 胎盤(placenta)や母乳(milk)を通じて供給される。
- 網膜色素変性症(retinitis pigmentosa)の患者ではDHAの血中濃度が低いと報告されています。
必須脂肪酸欠乏症(Essential Fatty Acid Deficiency):
- 非必須多価不飽和脂肪酸(nonessential polyenoic acids, ω9系)が、リン脂質、複合脂質、膜内で必須脂肪酸の代わりをします。
- 血漿脂質中のトリエン:テトラエン比(triene:tetraene ratio)は、必須脂肪酸欠乏症の程度を診断するために使用されます。
エイコサノイド(Eicosanoids)はC20多価不飽和脂肪酸から形成される
- アラキドン酸(arachidonate)および他のC20多価不飽和脂肪酸は、以下のエイコサノイド(eicosanoids)を生成:
- プロスタグランジン(prostaglandins, PG)
- トロンボキサン(thromboxanes, TX)
- ロイコトリエン(leukotrienes, LT)
- リポキシン(lipoxins, LX)
- 作用:
- 局所ホルモン(local hormones)として作用し、Gタンパク質共役受容体(G-protein–linked receptors)を介して生化学的効果を発揮。
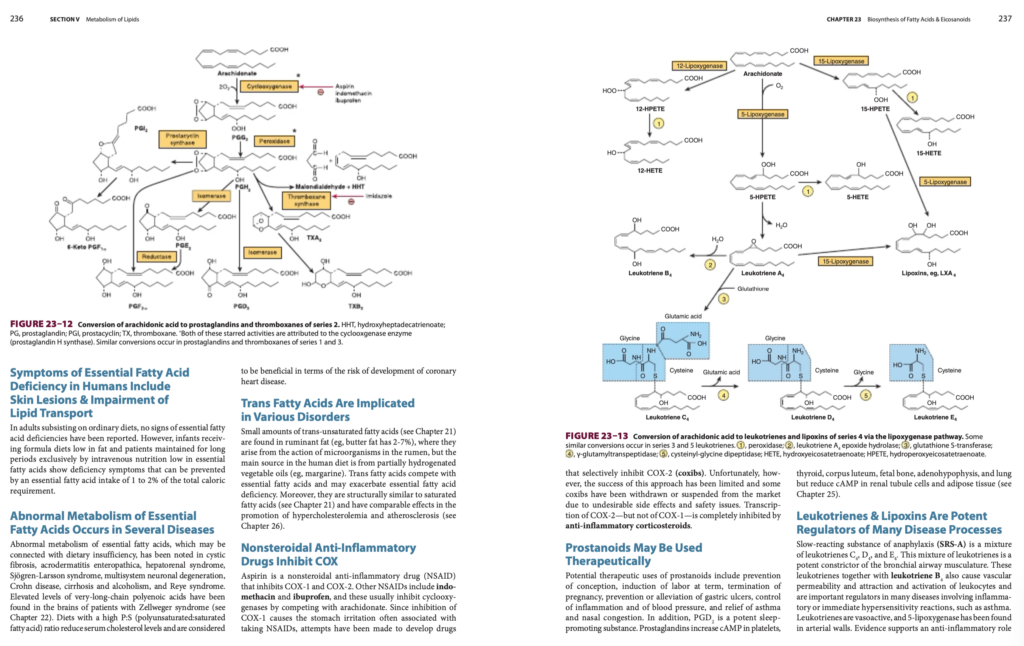
シクロオキシゲナーゼ経路(Cyclooxygenase Pathway)はプロスタノイド(Prostanoids)の合成を担う
- 経路:
- シクロオキシゲナーゼ(cyclooxygenase, COX)が触媒する最初の反応では、2分子の酸素が消費されます。
- COXには2つのアイソ酵素(isoenzymes)が存在:
- COX-1
- COX-2
- 生成物:エンドペルオキシド(endoperoxide, PGH)は、以下に変換:
- プロスタグランジンD(PGD)
- プロスタグランジンE(PGE)
- トロンボキサンA2(TXA2)
- プロスタサイクリン(PGI2)
- プロスタノイドの生理活性:
- トロンボキサン(thromboxanes, TXA2):
- 血小板(platelets)で合成され、血管収縮(vasoconstriction)と血小板凝集(platelet aggregation)を引き起こす。
- 低用量アスピリン(aspirin)によって合成が阻害。
- プロスタサイクリン(prostacyclin, PGI2):
- 血管壁で生成され、血小板凝集の強力な阻害剤。
- トロンボキサン(thromboxanes, TXA2):
ロイコトリエン(Leukotrienes)およびリポキシン(Lipoxins)はリポキシゲナーゼ経路(Lipoxygenase Pathway)で形成される
- ロイコトリエン:
- 白血球(leukocytes)、
肥満細胞(mastocytoma cells)、
血小板(platelets)、
マクロファージ(macrophages)
で生成。 - 5-リポキシゲナーゼ(5-lipoxygenase)によって作られる(図23–13参照)。
- 白血球(leukocytes)、
- リポキシン:
- 白血球で形成され、複数のリポキシゲナーゼの協働で生成。
臨床的側面(Clinical Aspects)
- 必須脂肪酸と疾患予防:
- 必須脂肪酸の摂取は、
心血管疾患(cardiovascular disease)、
がん(cancer)、
関節炎(arthritis)、
糖尿病(diabetes mellitus)、および
神経疾患(neurologic conditions)
の発症リスクを低下させることが示されています。 - メカニズム:
- 遺伝子発現の変化
- プロスタノイドの産生
- 膜の組成の変化
- 必須脂肪酸の摂取は、
必須脂肪酸欠乏症(Essential Fatty Acid Deficiency)の症状には皮膚病変(Skin Lesions)と脂質輸送の障害(Impairment of Lipid Transport)が含まれる
- 成人: 通常の食事を摂取している成人では、必須脂肪酸欠乏症の兆候は報告されていません。
- 乳幼児と長期静脈栄養の患者:
- 脂肪が少ない粉ミルク(formula diets)を摂取している乳幼児や、必須脂肪酸が不足した長期間の静脈栄養(intravenous nutrition)を受けている患者では、欠乏症状が見られます。
- 必須脂肪酸を総カロリー摂取量の1–2%摂取することで、これらの症状を予防可能です。
プロスタノイド(Prostanoids)の治療用途
- 治療的用途:
- 妊娠の予防(prevention of conception)
- 分娩誘発(induction of labor at term)
- 妊娠中絶(termination of pregnancy)
- 胃潰瘍の予防または緩和(prevention or alleviation of gastric ulcers)
- 炎症(inflammation)や血圧(blood pressure)の制御
- 喘息(asthma)および鼻詰まり(nasal congestion)の緩和
- PGD₂の特性:
- 強力な睡眠促進物質(sleep-promoting substance)。
- cAMPレベルへの影響:
- 血小板(platelets)、
甲状腺(thyroid)、
黄体(corpus luteum)、
胎児骨(fetal bone)、
下垂体前葉(adenohypophysis)、
肺(lung)
ではcAMPを増加させます。 - 腎細管細胞(renal tubule cells)および脂肪組織(adipose tissue)ではcAMPを減少させます(第25章参照)。
- 血小板(platelets)、
ロイコトリエン(Leukotrienes)とリポキシン(Lipoxins)は多くの疾患プロセスの調節因子
- ロイコトリエン:
- アナフィラキシー遅発反応物質(slow-reacting substance of anaphylaxis, SRS-A):
- ロイコトリエンC₄、D₄、E₄の混合物。
- 気管支平滑筋(bronchial airway musculature)の強力な収縮を引き起こします。
- 血管透過性(vascular permeability)の増加と白血球(leukocytes)の誘引および活性化を引き起こします。
- 喘息(asthma)や即時型過敏反応(immediate hypersensitivity reactions)を含む多くの炎症性疾患において重要。
- アナフィラキシー遅発反応物質(slow-reacting substance of anaphylaxis, SRS-A):
- リポキシン:
- 血管作動性(vasoactive)および免疫調節機能(immunoregulatory function)を持つ。
- 免疫応答の反規制化合物(counterregulatory compounds, chalones)として作用。
必須脂肪酸(Essential Fatty Acids)の異常代謝
必須脂肪酸の異常代謝は、以下の疾患で見られます:
- 嚢胞性線維症(cystic fibrosis)
- 腸性肢端皮膚炎(acrodermatitis enteropathica)
- 肝腎症候群(hepatorenal syndrome)
- シェーグレン・ラーション症候群(Sjögren-Larsson syndrome)
- 多系統神経変性(multisystem neuronal degeneration)
- クローン病(Crohn disease)
- 肝硬変(cirrhosis)およびアルコール依存症(alcoholism)
- ライ症候群(Reye syndrome)
例外:
- ゼルウェガー症候群(Zellweger syndrome, 第22章参照)患者の脳では、非常に長鎖の多価不飽和脂肪酸(very-long-chain polyenoic acids)の濃度が上昇しています。
高いP:S比(多価不飽和脂肪酸:飽和脂肪酸):
- 高P:S比の食事は血清コレステロール(serum cholesterol)を低下させ、冠動脈性心疾患(coronary heart disease)のリスクを減少させると考えられています。
トランス脂肪酸(Trans Fatty Acids)はさまざまな障害に関連
- トランス脂肪酸は反芻動物(ruminants)の脂肪(例:バター脂肪[butter fat])に少量含まれますが、人間の食事での主な供給源は部分水素添加植物油(partially hydrogenated vegetable oils, 例:マーガリン)です。
- 影響:
- 必須脂肪酸と競合し、必須脂肪酸欠乏症を悪化させる可能性があります。
- 構造的に飽和脂肪酸に似ており、同様に高コレステロール血症(hypercholesterolemia)や動脈硬化症(atherosclerosis)を促進します(第21章および第26章参照)。
非ステロイド性抗炎症薬(NSAIDs)はCOXを阻害する
- アスピリン(aspirin)は非ステロイド性抗炎症薬(nonsteroidal anti-inflammatory drug, NSAID)であり、COX-1およびCOX-2を阻害します。
- 他のNSAIDsにはインドメタシン(indomethacin)やイブプロフェン(ibuprofen)が含まれ、通常アラキドン酸(arachidonate)と競合してシクロオキシゲナーゼを阻害します。
COX-2特異的阻害薬(coxibs):
- 安全性の問題や副作用により、一部の薬剤は市場から撤回されています。
コルチコステロイド:
- COX-2の転写を完全に阻害する一方、COX-1には影響しません。

重要箇所穴埋め問題
Q1: Acetyl-CoA Carboxylase(アセチルCoAカルボキシラーゼ)
The enzyme Acetyl-CoA Carboxylase catalyzes the conversion of acetyl-CoA to _________, which is a key intermediate in fatty acid synthesis. This reaction requires _________ as a cofactor and _________ as an energy source.
Answer: malonyl-CoA / biotin / ATP
Explanation:
アセチルCoAカルボキシラーゼは、脂肪酸合成の律速段階 (rate-limiting step) であり、アセチルCoAをマロニルCoA (malonyl-CoA) に変換します。この反応には、ビオチン (biotin) が補因子 (cofactor) として必要であり、ATPがエネルギー源 (energy source) として必要です。
Q2: Allosteric and Hormonal Regulation(アロステリックおよびホルモン調節)
Acetyl-CoA Carboxylase is allosterically activated by _________ and inhibited by _________. Hormonally, insulin activates the enzyme through _________.
Answer: citrate / long-chain acyl-CoA / dephosphorylation
Explanation:
クエン酸 (citrate) はアロステリックにアセチルCoAカルボキシラーゼを活性化し、長鎖アシルCoA (long-chain acyl-CoA) はその活性を阻害します。インスリン (insulin) は、酵素を脱リン酸化 (dephosphorylation) することにより、活性を高めます。
Q3: Fatty Acid Synthase(脂肪酸合成酵素)
The Fatty Acid Synthase complex is a _________ that consists of _________ enzyme active sites along with an acyl carrier protein (ACP) on each polypeptide chain. The ACP contains a derivative of _________.
Answer: homodimer / 6 / pantothenic acid
Explanation:
脂肪酸合成酵素複合体 (Fatty Acid Synthase) はホモダイマー (homodimer) であり、各ペプチド鎖に6つの酵素活性部位 (enzyme active sites) を持ち、アシルキャリアタンパク質 (acyl carrier protein, ACP) も含まれています。ACPは、パントテン酸 (pantothenic acid) の誘導体を含んでいます。
Q4: Steps of Fatty Acid Synthesis(脂肪酸合成のステップ)
The elongation of fatty acid chains occurs through repeated cycles of _________, _________, and _________, with NADPH serving as the reducing equivalent.
Answer: condensation / reduction / dehydration
Explanation:
脂肪酸の鎖伸長は、縮合 (condensation)、還元 (reduction)、および脱水 (dehydration) という3つの主要な段階を繰り返すことで行われます。この過程では、NADPHが還元等価物 (reducing equivalent) として使用されます。
Q5: Sources of NADPH(NADPHの供給源)
The two major sources of NADPH for fatty acid synthesis are the _________ and the _________, which involve the enzymes glucose-6-phosphate dehydrogenase and malic enzyme, respectively.
Answer: pentose phosphate pathway / malic enzyme pathway
Explanation:
脂肪酸合成に必要なNADPHの主要な供給源は、ペントースリン酸経路 (pentose phosphate pathway) とリンゴ酸シャトル (malic enzyme pathway) です。これらの経路では、グルコース6-リン酸デヒドロゲナーゼ (glucose-6-phosphate dehydrogenase) とリンゴ酸酵素 (malic enzyme) が関与します。
Q6: Role of Malonyl-CoA(マロニルCoAの役割)
Malonyl-CoA not only serves as a precursor in fatty acid synthesis but also inhibits _________, thereby preventing the simultaneous occurrence of fatty acid _________ and _________.
Answer: carnitine palmitoyltransferase 1 (CPT1) / synthesis / oxidation
Explanation:
マロニルCoA (Malonyl-CoA) は、脂肪酸合成の前駆体 (precursor) であるだけでなく、カルニチンパルミトイルトランスフェラーゼ1 (Carnitine Palmitoyltransferase 1, CPT1) を阻害します。これにより、脂肪酸の合成 (synthesis) と酸化 (oxidation) が同時に行われるのを防ぎます。
Q7: Essential Fatty Acids(必須脂肪酸)
Humans cannot synthesize the essential fatty acids _________ and _________ because they lack the enzymes _________ and Δ15 desaturases.
Answer: linoleic acid / α-linolenic acid / Δ12 desaturases
Explanation:
ヒトは、リノール酸 (linoleic acid) およびα-リノレン酸 (α-linolenic acid) を体内で合成できません。これは、Δ12およびΔ15不飽和酵素 (desaturases) が欠如しているためであり、これらの必須脂肪酸 (essential fatty acids) は食事からの摂取が必要です。
Q8: Desaturation and Elongation of Fatty Acids(脂肪酸の不飽和化と伸長)
The enzyme _________ introduces double bonds into fatty acids, while the _________ enzymes are responsible for chain elongation. The first double bond is typically inserted at the _________ position.
Answer: desaturase / elongation / Δ9
Explanation:
デサチュラーゼ (desaturase) は脂肪酸に二重結合 (double bonds) を導入する酵素であり、エロンゲーション酵素 (elongation enzymes) は脂肪酸鎖の伸長 (chain elongation) に関与します。最初の二重結合は通常Δ9の位置に導入されます。
Q9: Eicosanoid Synthesis(エイコサノイド合成)
The cyclooxygenase pathway produces _________ and _________, while the lipoxygenase pathway produces _________, all of which play roles in inflammation and immune responses.
Answer: prostaglandins / thromboxanes / leukotrienes
Explanation:
シクロオキシゲナーゼ経路 (cyclooxygenase pathway) では、プロスタグランジン (prostaglandins) とトロンボキサン (thromboxanes) が生成され、リポキシゲナーゼ経路 (lipoxygenase pathway) ではロイコトリエン (leukotrienes) が生成されます。これらのエイコサノイド (eicosanoids) は、炎症 (inflammation) や免疫反応 (immune responses) に関与します。
*エイコサノイド:
プロスタグランジン(prostaglandins)、
トロンボキサン(thromboxanes)、
ロイコトリエン(leukotrienes)
リポキシン(lipoxins)
Q10: Regulation of Fatty Acid Synthesis(脂肪酸合成の調節)
A high-carbohydrate diet increases fatty acid synthesis via _________, while fasting inhibits it via the actions of _________ and _________.
Answer: insulin / glucagon / epinephrine
Explanation:
高炭水化物食 (high-carbohydrate diet) は、インスリン (insulin) を介して脂肪酸合成 (fatty acid synthesis) を促進します。一方、飢餓状態 (fasting) では、グルカゴン (glucagon) とエピネフリン (epinephrine) が脂肪酸合成を阻害します。
Q1: Fatty Acid Synthesis(脂肪酸合成)
Fatty acid synthesis occurs in the _________ of the cell and involves the conversion of _________ into _________ as the final product.
Answer: cytosol / acetyl-CoA / palmitate
Explanation:
脂肪酸合成 (fatty acid synthesis) は、細胞質 (cytosol) で行われ、アセチルCoA (acetyl-CoA) からパルミチン酸 (palmitate) への変換が最終生成物 (final product) です。
Q2: Cofactors in Fatty Acid Synthesis(脂肪酸合成に必要な補因子)
The synthesis of fatty acids requires several cofactors, including _________ as an energy source, _________ as a source of reducing equivalents, and _________ for carboxylation reactions.
Answer: ATP / NADPH / biotin
Explanation:
脂肪酸合成には、エネルギー源 (energy source) としてATP、還元等価物 (reducing equivalents) としてNADPH、カルボキシル化 (carboxylation) 反応のためのビオチン (biotin) が必要です。
Q3: Role of Malonyl-CoA(マロニルCoAの役割)
The formation of malonyl-CoA is the first and rate-limiting step in fatty acid synthesis, where _________ is carboxylated by the enzyme _________ using _________ as a cofactor.
Answer: acetyl-CoA / acetyl-CoA carboxylase / biotin
Explanation:
マロニルCoA (malonyl-CoA) の形成は、脂肪酸合成の最初かつ律速段階 (rate-limiting step) です。この反応では、アセチルCoA (acetyl-CoA) がアセチルCoAカルボキシラーゼ (acetyl-CoA carboxylase) によってカルボキシル化され、ビオチン (biotin) が補因子 (cofactor) として機能します。
Q4: Composition of Fatty Acid Synthase(脂肪酸合成酵素の構造)
Fatty Acid Synthase is a _________ with two identical polypeptide chains, each containing six enzyme active sites and an _________, which includes a derivative of _________.
Answer: homodimer / acyl carrier protein (ACP) / pantothenic acid
Explanation:
脂肪酸合成酵素 (fatty acid synthase) はホモダイマー (homodimer) であり、各ペプチド鎖に6つの酵素活性部位 (enzyme active sites) とアシルキャリアタンパク質 (acyl carrier protein, ACP) が含まれています。ACPは、パントテン酸 (pantothenic acid) の誘導体を含んでいます。
Q5: Essential Fatty Acids(必須脂肪酸)
Humans must obtain essential fatty acids like _________ and _________ from their diet because they lack the enzymes _________ and Δ15 desaturases needed to synthesize them.
Answer: linoleic acid / α-linolenic acid / Δ12 desaturase
Explanation:
ヒトは、リノール酸 (linoleic acid) およびα-リノレン酸 (α-linolenic acid) を食事から摂取する必要があります。これは、これらの必須脂肪酸 (essential fatty acids) を合成するためのΔ12およびΔ15デサチュラーゼ (desaturase) 酵素を欠いているためです。
Q6: Functions of Prostaglandins(プロスタグランジンの機能)
Prostaglandins are eicosanoids that regulate _________, mediate _________, and influence physiological processes like _________.
Answer: inflammation / pain / reproduction
Explanation:
プロスタグランジン (prostaglandins) は、炎症 (inflammation) を調節し、痛み (pain) を媒介し、さらに生殖 (reproduction) などの生理的プロセスにも影響を与えます。
Q7: NSAID Mechanism of Action(NSAIDの作用機序)
Nonsteroidal anti-inflammatory drugs (NSAIDs) such as _________ inhibit the synthesis of _________ by blocking the enzyme _________.
Answer: aspirin / prostaglandins / cyclooxygenase (COX)
Explanation:
非ステロイド性抗炎症薬 (NSAIDs) であるアスピリン (aspirin) は、シクロオキシゲナーゼ (cyclooxygenase, COX) を阻害することで、プロスタグランジン (prostaglandins) の合成をブロックします。
Q8: Role of Eicosanoids(エイコサノイドの役割)
Eicosanoids are derived from _________ and include bioactive lipids such as _________ and _________, which play key roles in inflammation and immune responses.
Answer: arachidonic acid / prostaglandins / leukotrienes
Explanation:
エイコサノイド (eicosanoids) は、アラキドン酸 (arachidonic acid) から生成される生理活性脂質であり、プロスタグランジン (prostaglandins) およびロイコトリエン (leukotrienes) を含みます。これらは炎症 (inflammation) や免疫応答 (immune responses) において重要な役割を果たします。
Q9: Tissue Sites of Fatty Acid Synthesis(脂肪酸合成の組織部位)
Fatty acid synthesis occurs in tissues such as the _________, _________, and _________, which are metabolically active in lipid metabolism.
Answer: liver / adipose tissue / lactating mammary gland
Explanation:
脂肪酸合成は、肝臓 (liver)、脂肪組織 (adipose tissue)、および授乳中の乳腺 (lactating mammary gland) で行われます。これらの組織は、脂質代謝において代謝的に活性な部位です。
Q10: Energy Requirements of Fatty Acid Synthesis(脂肪酸合成のエネルギー要求)
The synthesis of one molecule of palmitate requires _________ ATP, _________ NADPH, and _________ molecules of acetyl-CoA.
Answer: 7 / 14 / 8
Explanation:
1分子のパルミチン酸 (palmitate) を合成するためには、7分子のATP、14分子のNADPH、および8分子のアセチルCoA (acetyl-CoA) が必要です。
Q11: Role of NADPH in Fatty Acid Synthesis(脂肪酸合成におけるNADPHの役割)
NADPH required for fatty acid synthesis is supplied by the _________ and the _________ pathways, which are most active in tissues like the _________.
Answer: pentose phosphate / malic enzyme / liver
Explanation:
脂肪酸合成に必要なNADPHは、ペントースリン酸経路 (pentose phosphate pathway) およびマレック酵素 (malic enzyme) 経路から供給されます。これらの経路は、肝臓 (liver) などの活発な脂肪酸合成を行う組織で特に活性化されます。
Q1: Acetyl-CoA Transport(アセチルCoAの輸送)
Acetyl-CoA is produced in the _________, but since it cannot cross the _________ directly, it is converted to _________ for transport into the cytosol.
Answer: mitochondrial matrix / mitochondrial membrane / citrate
Explanation:
アセチルCoA (acetyl-CoA) はミトコンドリアマトリックス (mitochondrial matrix) で生成されますが、ミトコンドリア膜 (mitochondrial membrane) を直接通過できません。そのため、クエン酸 (citrate) に変換され、細胞質 (cytosol) に輸送されます。
Q2: Citrate Transport System(クエン酸輸送システム)
The transport of citrate from mitochondria to the cytosol requires the _________ transporter, where citrate is cleaved by _________ to form acetyl-CoA and _________ in the cytosol.
Answer: tricarboxylate / ATP-citrate lyase / oxaloacetate
Explanation:
クエン酸は三カルボン酸トランスポーター (tricarboxylate transporter) を介してミトコンドリアから細胞質へ輸送されます。細胞質では、ATP-クエン酸リアーゼ (ATP-citrate lyase) によってクエン酸がアセチルCoA (acetyl-CoA) とオキサロ酢酸 (oxaloacetate) に分解されます。
Q3: Role of Malate(マレートの役割)
After the breakdown of citrate in the cytosol, oxaloacetate is converted to _________ by _________, and this product can generate _________, which is essential for fatty acid synthesis.
Answer: malate / malate dehydrogenase / NADPH
Explanation:
細胞質でのクエン酸の分解により生成されるオキサロ酢酸 (oxaloacetate) は、マレートデヒドロゲナーゼ (malate dehydrogenase) によってマレート (malate) に変換されます。その後、マレック酵素 (malic enzyme) によってピルビン酸 (pyruvate) に変換される際、NADPHが生成されます。このNADPHは、脂肪酸合成に必要な還元当量 (reducing equivalents) として機能します。
Q4: Regulation of Acetyl-CoA Carboxylase(アセチルCoAカルボキシラーゼの調節)
Acetyl-CoA carboxylase is activated by _________ and _________, while it is inhibited by _________ through negative feedback.
Answer: citrate / dephosphorylation / long-chain acyl-CoA
Explanation:
アセチルCoAカルボキシラーゼ (acetyl-CoA carboxylase) は、クエン酸 (citrate) によってアロステリックに活性化され、脱リン酸化 (dephosphorylation) によっても活性化されます。一方、長鎖アシルCoA (long-chain acyl-CoA) による負のフィードバック (negative feedback) によって阻害されます。

Q5: Hormonal Regulation of Fatty Acid Synthesis(脂肪酸合成のホルモン調節)
Fatty acid synthesis is promoted by _________ through the dephosphorylation of acetyl-CoA carboxylase, while it is inhibited by _________ and _________ through enzyme phosphorylation.
Answer: insulin / glucagon / epinephrine
Explanation:
脂肪酸合成 (fatty acid synthesis) は、インスリン (insulin) によるアセチルCoAカルボキシラーゼ (acetyl-CoA carboxylase) の脱リン酸化 (dephosphorylation) によって促進されます。一方、グルカゴン (glucagon) およびエピネフリン (epinephrine) によるリン酸化 (phosphorylation) によって抑制されます。
Q6: Fatty Acid Elongation(脂肪酸の伸長)
Fatty acid chain elongation occurs in the _________ and involves the use of _________ as a two-carbon donor and _________ as a reducing agent.
Answer: endoplasmic reticulum / malonyl-CoA / NADPH
Explanation:
脂肪酸の伸長 (elongation) は、小胞体 (endoplasmic reticulum) で行われ、2炭素供与体 (two-carbon donor) としてマロニルCoA (malonyl-CoA) が使用されます。NADPHは、還元剤 (reducing agent) として機能します。
Q7: Regulation of Lipogenesis(脂肪生成の調節)
Lipogenesis is increased by a _________ diet, while it is decreased during _________ and under conditions of _________ deficiency.
Answer: high-carbohydrate / fasting / insulin
Explanation:
脂肪生成 (lipogenesis) は、高炭水化物食 (high-carbohydrate diet) によって促進され、飢餓 (fasting) 状態およびインスリン欠乏 (insulin deficiency) によって抑制されます。
Q8: Role of Insulin in Fatty Acid Synthesis(脂肪酸合成におけるインスリンの役割)
Insulin increases fatty acid synthesis by promoting _________ entry into cells, activating _________, and reducing levels of circulating _________.
Answer: glucose / acetyl-CoA carboxylase / free fatty acids
Explanation:
インスリン (insulin) は、細胞へのグルコース (glucose) の取り込みを促進し、アセチルCoAカルボキシラーゼ (acetyl-CoA carboxylase) を活性化し、血漿遊離脂肪酸 (free fatty acids) の濃度を低下させることで、脂肪酸合成 (fatty acid synthesis) を促進します。
Q9: Regulation by Acyl-CoA(アシルCoAによる調節)
Long-chain acyl-CoA inhibits _________, _________, and the transport of _________ from mitochondria to the cytosol.
Answer: acetyl-CoA carboxylase / pyruvate dehydrogenase / citrate
Explanation:
長鎖アシルCoA (long-chain acyl-CoA) は、アセチルCoAカルボキシラーゼ (acetyl-CoA carboxylase) およびピルビン酸デヒドロゲナーゼ (pyruvate dehydrogenase) を抑制し、クエン酸 (citrate) のミトコンドリアから細胞質への輸送も阻害します。
Q10: Role of ATP in Fatty Acid Synthesis(脂肪酸合成におけるATPの役割)
ATP is required for the activity of _________, the formation of _________, and the cleavage of _________ in the cytosol.
Answer: ATP-citrate lyase / malonyl-CoA / citrate
Explanation:
ATPは、ATP-クエン酸リアーゼ (ATP-citrate lyase) の活性、マロニルCoA (malonyl-CoA) の形成、および細胞質でのクエン酸 (citrate) の分解に必要です。
Q1: Essential Fatty Acids(必須脂肪酸)
Essential fatty acids, including _________ and _________, cannot be synthesized by mammals because they lack the enzymes _________ and Δ15 desaturase.
Answer: linoleic acid / α-linolenic acid / Δ12 desaturase
Explanation:
リノール酸 (linoleic acid) およびα-リノレン酸 (α-linolenic acid) は必須脂肪酸 (essential fatty acids) であり、哺乳類はΔ12およびΔ15不飽和酵素 (desaturase enzymes) を持たないため、これらの脂肪酸を合成することができません。
Q2: Polyunsaturated Fatty Acids(多価不飽和脂肪酸)
Polyunsaturated fatty acids (PUFAs) are formed by the action of _________, which introduces double bonds, and _________, which increases chain length. These enzymes are located in the _________.
Answer: desaturase / elongase / endoplasmic reticulum
Explanation:
多価不飽和脂肪酸 (polyunsaturated fatty acids) は、デサチュラーゼ (desaturase) による二重結合の導入と、エロンゲース (elongase) による鎖の伸長によって生成されます。これらの酵素は小胞体 (endoplasmic reticulum) に存在します。
Q3: Synthesis of Monounsaturated Fatty Acids(一価不飽和脂肪酸の合成)
Monounsaturated fatty acids (MUFAs) are synthesized from _________ by the action of the _________ system, which introduces a double bond at the _________ position.
Answer: saturated fatty acids / Δ9 desaturase / Δ9
Explanation:
一価不飽和脂肪酸 (monounsaturated fatty acids) は、飽和脂肪酸 (saturated fatty acids) からΔ9デサチュラーゼ (Δ9 desaturase) 系の作用によって生成されます。この酵素は、鎖内のΔ9位置に二重結合を導入します。
Q4: Desaturation of Fatty Acids(脂肪酸の不飽和化)
Animals can introduce double bonds at positions _________, _________, and _________, but they lack the ability to introduce double bonds beyond Δ9.
Answer: Δ4 / Δ5 / Δ6
Explanation:
動物は、Δ4、Δ5、およびΔ6の位置に二重結合を導入する能力を持っていますが、Δ9を超える位置には二重結合を導入する能力がありません。これが、必須脂肪酸 (essential fatty acids) を食事から摂取しなければならない理由の一つです。
Q5: Essential Fatty Acids and Health(必須脂肪酸と健康)
Essential fatty acids play a role in the formation of _________, _________, and _________, which are critical for inflammation, immune response, and cardiovascular health.
Answer: prostaglandins / thromboxanes / leukotrienes
Explanation:
必須脂肪酸 (essential fatty acids) は、プロスタグランジン (prostaglandins)、トロンボキサン (thromboxanes)、およびロイコトリエン (leukotrienes) の形成に関与します。これらの分子は、炎症 (inflammation)、免疫応答 (immune response)、および心血管系の健康 (cardiovascular health) に重要な役割を果たします。
Q6: Structure and Function of Eicosanoids(エイコサノイドの構造と機能)
Eicosanoids are derived from _________ and include _________, _________, and leukotrienes, which act as local hormones to mediate inflammation.
Answer: arachidonic acid / prostaglandins / thromboxanes
Explanation:
エイコサノイド (eicosanoids) は、アラキドン酸 (arachidonic acid) から生成される分子群であり、プロスタグランジン (prostaglandins)、トロンボキサン (thromboxanes)、およびロイコトリエン (leukotrienes) を含みます。これらは局所ホルモン (local hormones) として機能し、炎症 (inflammation) を媒介します。
Q7: Cyclooxygenase Pathway(シクロオキシゲナーゼ経路)
The cyclooxygenase (COX) pathway involves the enzyme _________, which converts arachidonic acid into _________, a precursor for _________ and thromboxanes.
Answer: cyclooxygenase / endoperoxide (PGH) / prostaglandins
Explanation:
シクロオキシゲナーゼ経路 (cyclooxygenase pathway) では、アラキドン酸 (arachidonic acid) がシクロオキシゲナーゼ (cyclooxygenase, COX) によってエンドペルオキシド (endoperoxide, PGH) に変換されます。この生成物は、プロスタグランジン (prostaglandins) およびトロンボキサン (thromboxanes) の前駆体 (precursor) となります。
Q8: Role of Prostaglandins(プロスタグランジンの役割)
Prostaglandins are synthesized through the cyclooxygenase pathway and play a role in _________, _________, and the regulation of _________.
Answer: inflammation / pain / blood coagulation
Explanation:
プロスタグランジン (prostaglandins) は、シクロオキシゲナーゼ (cyclooxygenase) 経路を通じて合成され、炎症 (inflammation)、痛み (pain)、および血液凝固 (blood coagulation) の調節に関与します。
Q9: Role of Δ9 Desaturase(Δ9デサチュラーゼの役割)
The Δ9 desaturase system introduces a double bond in _________ and _________ at the Δ9 position, producing _________ and oleoyl-CoA, respectively.
Answer: palmitoyl-CoA / stearoyl-CoA / palmitoleoyl-CoA
Explanation:
Δ9デサチュラーゼ (Δ9 desaturase) 系は、パルミトイルCoA (palmitoyl-CoA) およびステアロイルCoA (stearoyl-CoA) にΔ9位置で二重結合を導入し、それぞれパルミトレオイルCoA (palmitoleoyl-CoA) およびオレオイルCoA (oleoyl-CoA) を生成します。
Q10: Role of Lipoxygenase(リポキシゲナーゼの役割)
Lipoxygenase enzymes catalyze the formation of _________ and _________, which play key roles in _________ and immune responses.
Answer: leukotrienes / lipoxins / allergic reactions
Explanation:
リポキシゲナーゼ (lipoxygenase) 酵素は、ロイコトリエン (leukotrienes) およびリポキシン (lipoxins) の形成を触媒します。これらは、アレルギー反応 (allergic reactions) および免疫応答 (immune responses) に重要な役割を果たします。
Q1: Essential Fatty Acid Deficiency(必須脂肪酸欠乏症)
Essential fatty acid deficiency can result in _________, _________, and impairment of _________, especially in infants or patients on long-term intravenous nutrition.
Answer: skin lesions / growth retardation / lipid transport
Explanation:
必須脂肪酸欠乏症 (essential fatty acid deficiency) は、皮膚病変 (skin lesions)、成長遅延 (growth retardation)、および脂質輸送の障害 (impairment of lipid transport) を引き起こします。これらの症状は、特に脂肪の少ない粉ミルク (low-fat formula) を与えられた乳幼児や、長期間の静脈栄養 (intravenous nutrition) を受けている患者で観察されます。
Q2: Role of Leukotrienes(ロイコトリエンの役割)
Leukotrienes are derived from _________, and they function as _________, causing _________ in the bronchial airway musculature.
Answer: arachidonic acid / slow-reacting substances of anaphylaxis (SRS-A) / contraction
Explanation:
ロイコトリエン (leukotrienes) は、アラキドン酸 (arachidonic acid) から生成され、アナフィラキシー遅発反応物質 (slow-reacting substances of anaphylaxis, SRS-A) として機能します。これにより、気道平滑筋 (bronchial airway musculature) の収縮 (contraction) が引き起こされます。
Q3: Therapeutic Uses of Prostanoids(プロスタノイドの治療的用途)
Prostanoids can be used for the _________, _________, and _________, demonstrating their importance in reproductive and gastrointestinal health.
Answer: induction of labor / prevention of conception / prevention of gastric ulcers
Explanation:
プロスタノイド (prostanoids) は、分娩誘発 (induction of labor)、妊娠の予防 (prevention of conception)、および胃潰瘍の予防 (prevention of gastric ulcers) に使用されます。これらの治療的用途は、プロスタノイドが生殖および消化器系の健康において重要な役割を果たしていることを示しています。
Q4: Role of NSAIDs(NSAIDsの役割)
Nonsteroidal anti-inflammatory drugs (NSAIDs) like _________ inhibit _________ and prevent the synthesis of _________, thereby reducing pain and inflammation.
Answer: aspirin / cyclooxygenase (COX) / prostaglandins
Explanation:
非ステロイド性抗炎症薬 (NSAIDs) の一つであるアスピリン (aspirin) は、シクロオキシゲナーゼ (cyclooxygenase, COX) を阻害し、プロスタグランジン (prostaglandins) の合成を妨げます。これにより、痛み (pain) と炎症 (inflammation) が軽減されます。
Q5: Function of Lipoxins(リポキシンの機能)
Lipoxins are eicosanoids with _________, _________, and _________ functions, making them key players in immune response regulation.
Answer: vasoactive / anti-inflammatory / immunoregulatory
Explanation:
リポキシン (lipoxins) は、血管作動性 (vasoactive)、抗炎症 (anti-inflammatory)、および免疫調節 (immunoregulatory) の機能を持つエイコサノイド (eicosanoids) です。これらの特性により、免疫応答の調節において重要な役割を果たします。
Q6: Abnormal Metabolism of Essential Fatty Acids(必須脂肪酸の異常代謝)
Abnormal metabolism of essential fatty acids is associated with _________, _________, and _________, leading to metabolic and neurological disorders.
Answer: cystic fibrosis / Crohn’s disease / Sjögren-Larsson syndrome
Explanation:
必須脂肪酸の異常代謝 (abnormal metabolism of essential fatty acids) は、嚢胞性線維症 (cystic fibrosis)、クローン病 (Crohn’s disease)、およびシェーグレン・ラーション症候群 (Sjögren-Larsson syndrome) に関連しています。これらの状態は、代謝的および神経学的障害 (metabolic and neurological disorders) を引き起こす可能性があります。
Q7: Impact of Trans Fatty Acids(トランス脂肪酸の影響)
Trans fatty acids are found in _________ and _________, and they contribute to _________ by competing with essential fatty acids.
Answer: ruminant fat / partially hydrogenated vegetable oils / essential fatty acid deficiency
Explanation:
トランス脂肪酸 (trans fatty acids) は、反芻動物の脂肪 (ruminant fat) および部分水素添加植物油 (partially hydrogenated vegetable oils) に含まれています。これらは必須脂肪酸 (essential fatty acids) と競合し、必須脂肪酸欠乏症 (essential fatty acid deficiency) を悪化させる可能性があります。
Q8: Symptoms of Essential Fatty Acid Deficiency(必須脂肪酸欠乏症の症状)
Symptoms of essential fatty acid deficiency include _________, _________, and _________, particularly in infants on low-fat diets or long-term intravenous nutrition.
Answer: dermatitis / hair loss / impaired wound healing
Explanation:
必須脂肪酸欠乏症 (essential fatty acid deficiency) の症状には、皮膚炎 (dermatitis)、脱毛 (hair loss)、および創傷治癒の遅延 (impaired wound healing) が含まれます。これらの症状は、脂肪の少ない食事を摂取している乳幼児や、長期間の静脈栄養を受けている患者で特に顕著です。
Q9: Role of Eicosanoids(エイコサノイドの役割)
Eicosanoids are local hormones derived from _________ and include _________ and _________, which are key regulators of inflammation and immunity.
Answer: arachidonic acid / prostaglandins / leukotrienes
Explanation:
エイコサノイド (eicosanoids) は、アラキドン酸 (arachidonic acid) から生成される局所ホルモン (local hormones) であり、プロスタグランジン (prostaglandins) およびロイコトリエン (leukotrienes) を含みます。これらは、炎症 (inflammation) および免疫 (immunity) の主要な調節因子です。
Q10: Role of P:S Ratio(P:S比の役割)
A diet with a high polyunsaturated to saturated fat ratio (P:S ratio) is linked to reduced _________, decreased risk of _________, and improved _________ health.
Answer: serum cholesterol / coronary heart disease / cardiovascular
Explanation:
多価不飽和脂肪酸:飽和脂肪酸比 (polyunsaturated to saturated fat ratio, P:S ratio) が高い食事は、血清コレステロール (serum cholesterol) の低下、冠動脈性心疾患 (coronary heart disease) のリスク低下、および心血管の健康 (cardiovascular health) の改善に関連しています。

コメント