
Contents
- 1 呼吸中枢 (Respiratory Center)
- 2 背側呼吸群の機能 (Dorsal Respiratory Group)
- 3 呼吸調節中枢 (Pneumotaxic Center) の役割
- 4 腹側呼吸群 (Ventral Respiratory Group) の機能
- 5 肺膨張信号による吸息制限—ヘーリング・ブロイヤー反射 (Hering-Breuer Inflation Reflex)
- 6 全体的な呼吸中枢活動の調節 (Control of Overall Respiratory Center Activity)
- 7 呼吸の化学的調節 (Chemical Control of Respiration)
- 8 血液中のPco2 と H+ 濃度が肺換気に及ぼす定量的影響 (Quantitative Effects of Blood Pco2 and H+ Concentration on Alveolar Ventilation)
- 9 酸素の直接的影響 (Direct Effect of Oxygen)
- 10 末梢化学受容体系 (Peripheral Chemoreceptor System) – 酸素の呼吸制御における役割 (Role of Oxygen in Respiratory Control)
- 11 酸素不足による化学受容体の刺激 (Decreased Arterial Oxygen Stimulates the Chemoreceptors)
- 12 酸素欠乏による化学受容体の基本メカニズム (Basic Mechanism of Stimulation of the Chemoreceptors by O2 Deficiency)
- 13 CO2 と H+ の化学受容体への刺激 (Increased CO2 and H+ Concentration Stimulates the Chemoreceptors)
- 14 低酸素の慢性的な呼吸が呼吸をさらに刺激する—「順応現象」 (Chronic Breathing of Low O2 Stimulates Respiration Even More—The Phenomenon of “Acclimatization”)
- 15 Pco2、pH、Po2 の複合的な影響 (Composite Effects of Pco2, pH, and Po2 on Alveolar Ventilation)
- 16 運動中の呼吸調節 (Regulation of Respiration During Exercise)
- 17 呼吸に影響を与えるその他の要因 (Other Factors That Affect Respiration)
- 18 その他の呼吸に影響を与える要因
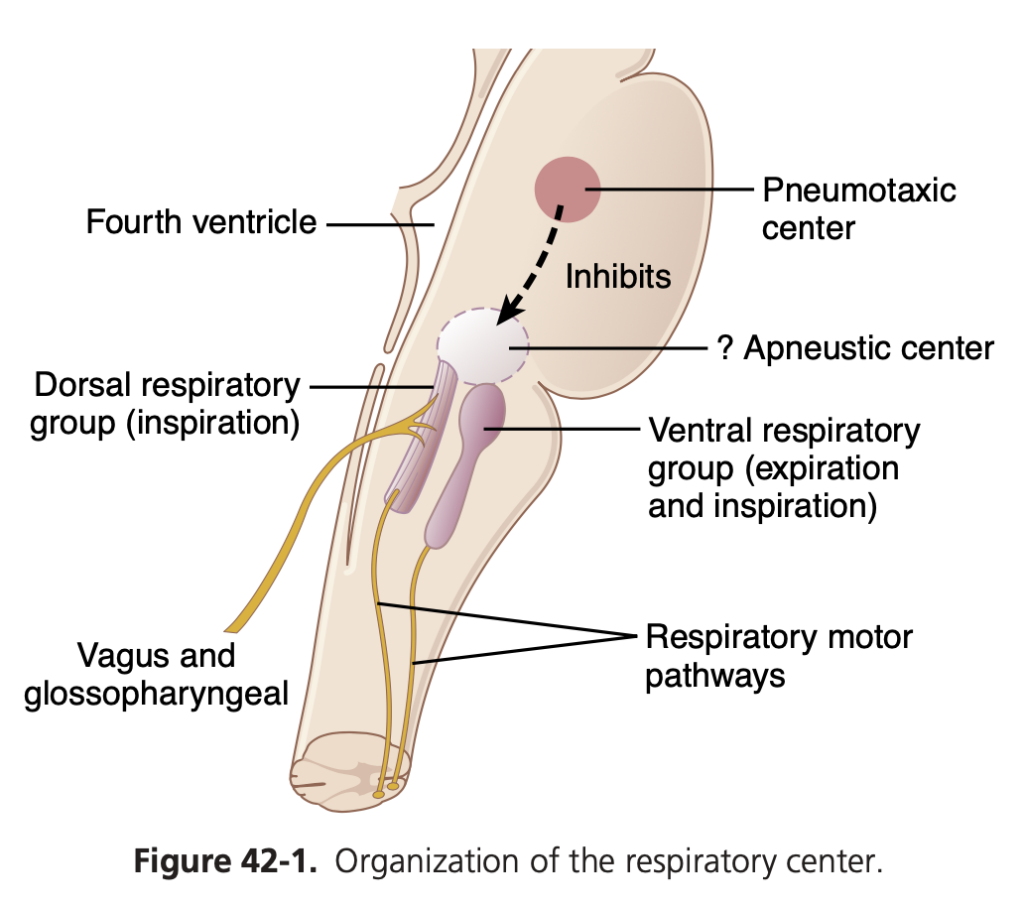
呼吸中枢 (Respiratory Center)
呼吸中枢は、脳幹の延髄 (medulla oblongata) と橋 (pons) に位置するいくつかの神経群で構成されています。以下の三つの主要な神経群に分かれています:
- 背側呼吸群 (Dorsal Respiratory Group): 延髄の背側部分に位置し、主に吸息 (inspiration) を引き起こします。
- 腹側呼吸群 (Ventral Respiratory Group): 延髄の腹側外側部分に位置し、主に呼息 (expiration) を引き起こします。
- 呼吸調節中枢 (Pneumotaxic Center): 橋の上部背側に位置し、主に呼吸の速度 (rate) と深さ (depth) を調節します。
背側呼吸群の機能 (Dorsal Respiratory Group)
背側呼吸群は呼吸の制御において重要な役割を果たし、延髄のほとんどの長さにわたっています。この神経群の大部分はトラクトスソリタリウス核 (Nucleus of the Tractus Solitarius, NTS) に位置しています。NTSは以下の感覚信号を受け取ります:
- 末梢化学受容体 (Peripheral Chemoreceptors)
- バロレセプター (Baroreceptors)
- 肝臓、膵臓、消化管の各部位の受容体 (Receptors in the Liver, Pancreas, and Gastrointestinal Tract)
- 肺内のさまざまな受容体 (Receptors in the Lungs)
吸息のリズム生成 (Rhythmical Inspiratory Discharges)
背側呼吸群は呼吸の基本的なリズムを生成します。すべての末梢神経が切断され、脳幹が延髄の上と下で切断されても、この神経群はまだ繰り返し吸息の神経活動電位を発生させます。このリズムの基本的な原因は不明ですが、原始的な動物では、ある神経群の活動が別の神経群を興奮させ、さらにその神経群が最初の神経群を抑制するような神経ネットワークが存在します。類似のネットワークが人間にも存在し、延髄内の背側呼吸群や隣接する領域が関与していると考えられています。
吸息の「ランプ」信号 (Inspiratory “Ramp” Signal)
吸息筋、主に横隔膜 (diaphragm) に送られる神経信号は、一瞬で強くなるものではなく、弱く始まり、約2秒間にわたって段階的に増加します。その後、信号は突然終了し、約3秒間の間に呼息が起こります。この周期が繰り返され、吸息信号が再び始まります。この「ランプ」信号の利点は、吸息中に肺の容量が安定して増加することです。
ランプ信号の2つの調節 (Two Qualities of the Inspiratory Ramp)
- ランプ信号の増加速度の調節 (Control of the Rate of Increase of the Ramp Signal): 激しい呼吸の時には、ランプが迅速に増加し、肺が速く満たされるようにします。
- ランプ信号の終了点の調節 (Control of the Limiting Point): ランプが早く終了すると吸息の持続時間が短くなり、呼息の持続時間も短くなります。この方法で呼吸の頻度 (frequency) が増加します。
←背側 腹側→
呼吸調節中枢 (Pneumotaxic Center) の役割
呼吸調節中枢 (Pneumotaxic Center) は、上部橋 (upper pons) のパラブラキア核 (nucleus parabrachialis) に位置し、吸息エリア (inspiratory area) に信号を送ります。この中枢の主要な役割は、吸息ランプ (inspiratory ramp) の「スイッチオフ」ポイントを制御することです。これにより、肺のサイクリングの充填相 (filling phase) の持続時間が調節されます。呼吸調節中枢からの信号が強いと、吸息はわずか0.5秒しか続かず、肺はわずかに膨らむだけです。信号が弱いと、吸息は5秒以上続き、肺はより多くの空気で満たされます。
呼吸調節中枢の機能 (Function of the Pneumotaxic Center)
呼吸調節中枢の機能は主に吸息を制限することです。この制限は、呼吸の頻度 (rate) を増加させる副次的な効果があります。吸息が制限されると呼息も短くなり、呼吸全体の周期が短くなります。強い呼吸調節中枢の信号は、呼吸頻度を30〜40回/分に増加させることができます。一方で、信号が弱いと呼吸頻度は3〜5回/分に減少します。
腹側呼吸群 (Ventral Respiratory Group) の機能
腹側呼吸群 (Ventral Respiratory Group) は、延髄の両側、背側呼吸群の神経群の約5ミリメートル前方および外側に位置しています。この神経群の機能は、背側呼吸群とはいくつかの重要な点で異なります:
- 通常の静かな呼吸中の不活性 (Inactivity During Normal Quiet Breathing): 腹側呼吸群の神経は、通常の静かな呼吸中にはほとんど活動しません。静かな呼吸は、背側呼吸群からの繰り返しの吸息信号によって引き起こされ、呼息は肺や胸郭の弾性反動 (elastic recoil) によって起こります。
- 基本的リズムの不参加 (Lack of Participation in Basic Rhythmic Oscillation): 腹側呼吸群の神経は、呼吸を制御する基本的なリズムには関与していません。
- 呼吸駆動の増加 (Increased Respiratory Drive): 呼吸駆動が通常より強くなると、背側呼吸エリアの基本的な振動メカニズムから腹側呼吸群に信号が伝わり、腹側呼吸エリアも追加の呼吸駆動に寄与します。
- 刺激による吸息と呼息 (Stimulation-Induced Inspiration and Expiration): 腹側呼吸群の神経の一部に電気刺激を与えると吸息が引き起こされ、別の神経に刺激を与えると呼息が引き起こされます。したがって、これらの神経群は吸息と呼息の両方に寄与し、特に強い呼息を必要とする際には腹部の筋肉に対して強力な信号を提供します。このため、非常に激しい運動中には過剰な呼吸駆動メカニズムとして機能します。
肺膨張信号による吸息制限—ヘーリング・ブロイヤー反射 (Hering-Breuer Inflation Reflex)
脳幹内の中枢神経系の呼吸調節メカニズムに加えて、肺からの感覚神経信号も呼吸の制御に役立ちます。最も重要なのは、気管支および細気管支 (bronchi and bronchioles) の筋肉部分に位置する伸展受容体 (stretch receptors) です。これらの受容体は、肺が過膨張 (overstretched) すると信号を迷走神経 (vagus nerve) を介して背側呼吸群に送ります。これにより、肺が過膨張すると吸息ランプが「スイッチオフ」され、さらに吸息が停止します。このメカニズムはヘーリング・ブロイヤー反射 (Hering-Breuer Inflation Reflex) と呼ばれ、呼吸調節中枢からの信号と同様に呼吸頻度を増加させます。
人間では、ヘーリング・ブロイヤー反射は潮気量 (tidal volume) が通常の3倍以上 (>≈1.5 L/呼吸) にならないと活性化されないため、通常の呼吸調節にはあまり寄与せず、過剰な肺膨張を防ぐための保護メカニズムとして機能しています。
全体的な呼吸中枢活動の調節 (Control of Overall Respiratory Center Activity)
ここまでで、吸息と呼息を引き起こす基本的なメカニズムについて説明しましたが、呼吸制御信号の強度がどのように増減するかを理解することも重要です。例えば、激しい運動中には酸素 (O2) の使用量と二酸化炭素 (CO2) の生成量が通常の20倍に増加することがあり、それに伴い肺の換気 (pulmonary ventilation) も増加する必要があります。この章の残りの部分では、体の呼吸ニーズに応じた換気の調節について説明します。
呼吸の化学的調節 (Chemical Control of Respiration)
呼吸の最終的な目標は、組織内の酸素 (O2)、二酸化炭素 (CO2)、および水素イオン (H+) の適切な濃度を維持することです。幸いなことに、呼吸活動はこれらの物質の変化に非常に敏感に反応します。
CO2 と H+ による呼吸中枢の直接的制御 (Direct Control of Respiratory Center Activity by CO2 and H+)
- 化学感受性領域 (Chemosensitive Area): 延髄の腹側表面 (ventral surface) の下に存在する化学感受性領域は、血液中のCO2やH+の変化に非常に敏感です。この領域は、背側呼吸群 (dorsal respiratory group) や腹側呼吸群 (ventral respiratory group) など他の呼吸中枢部分を興奮させます。
- H+ による化学感受性ニューロンの興奮 (Excitation of the Chemosensitive Neurons by H+): 化学感受性領域のセンサーニューロンは、特にH+ によって興奮します。H+ が直接的な重要な刺激であると考えられていますが、H+ イオンは血液脳関門 (blood-brain barrier) を通過しにくいため、血液中のH+ 濃度の変化はCO2 よりも刺激効果が小さいです。
- CO2 の間接的な刺激作用 (CO2 Indirectly Stimulates the Chemosensitive Neurons): CO2 は直接的な刺激効果は少ないものの、間接的に強力な効果を持っています。CO2 は組織の水と反応して炭酸 (carbonic acid) を形成し、これがH+ とHCO3− に解離します。H+ が強力な直接的刺激効果を持ちます。
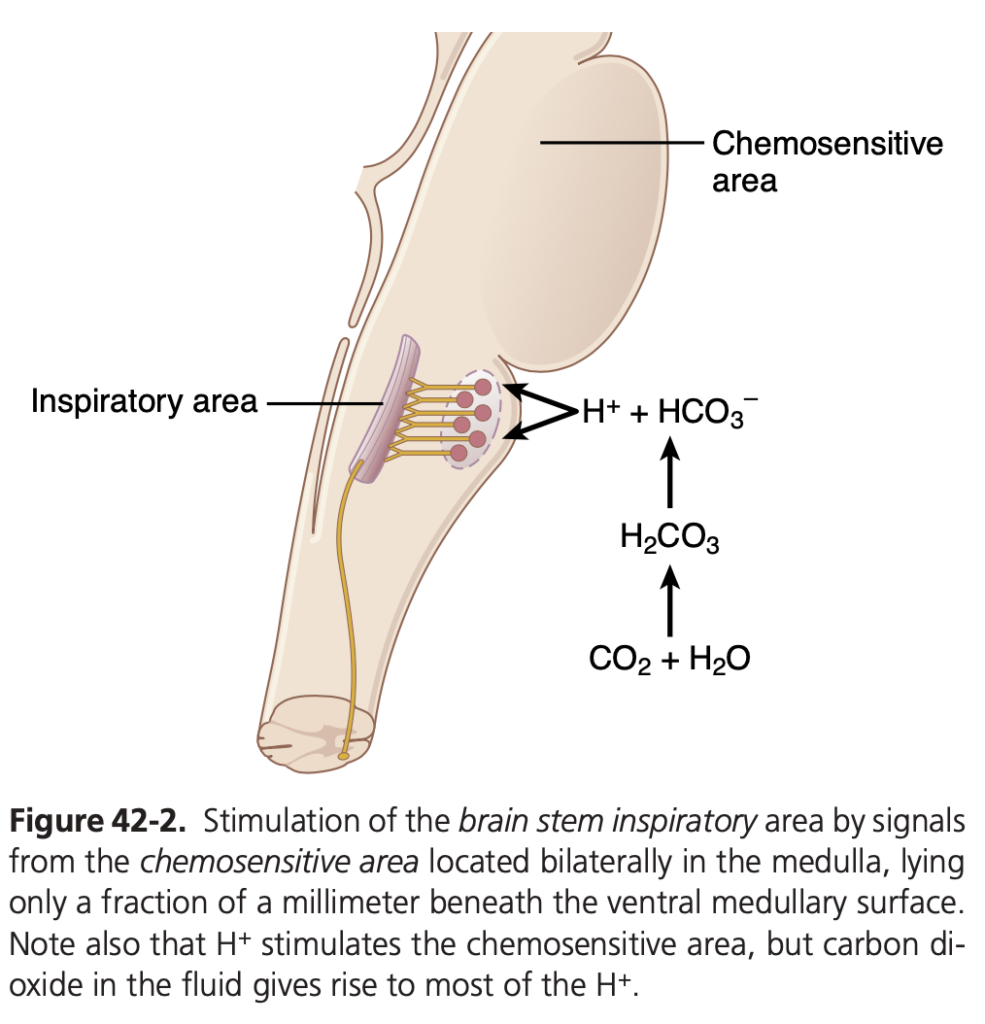
なぜ血液中のCO2 はH+ よりも強力な刺激を与えるのか? (Why CO2 is More Potent than H+)
血液脳関門はH+ にはあまり透過性がありませんが、CO2 はほぼその関門が存在しないかのように通過します。そのため、血液中のCO2 濃度が増加すると、延髄の間質液 (interstitial fluid) や脳脊髄液 (cerebrospinal fluid) のCO2 濃度も増加します。これにより、CO2 が水と反応して新たにH+ が生成されます。このため、血液中のCO2 濃度が増加すると、H+ 濃度も増加し、呼吸中枢の活動が強くなるのです。
CO2 の刺激効果の減衰 (Attenuated Stimulatory Effect of CO2)
- 初期の強い刺激効果 (Initial Strong Stimulatory Effect): 血液中のCO2 濃度が増加した最初の数時間は、呼吸中枢の興奮が非常に強いです。しかし、次第にその効果は1〜2日間で減少し、初期の約5分の1まで低下します。
- 腎臓による調整 (Renal Readjustment): CO2 がH+ 濃度を増加させた後、腎臓は血液中のHCO3− 濃度を増加させることで、H+ と結合しその濃度を減少させます。さらに、HCO3− は血液脳関門や血液脳脊髄液関門を通過してH+ と直接結合し、H+ 濃度を正常に戻します。これにより、CO2 の変化は急性には強力な影響を与えますが、数日後には慢性的な影響は弱まります。
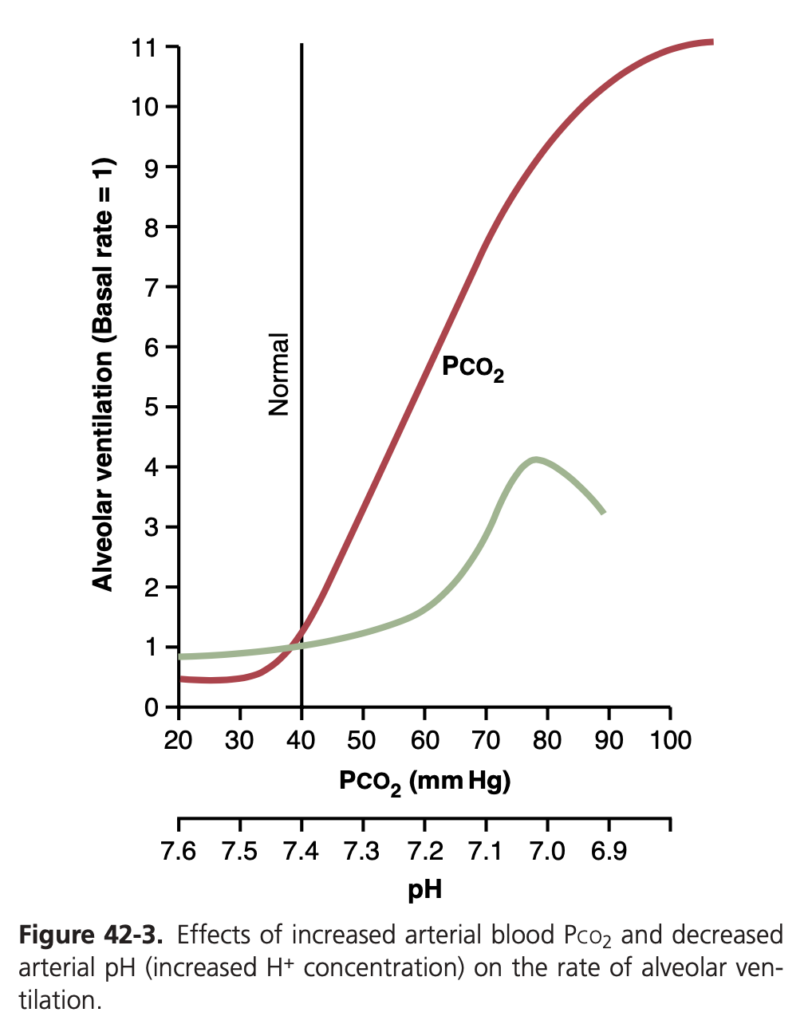
血液中のPco2 と H+ 濃度が肺換気に及ぼす定量的影響 (Quantitative Effects of Blood Pco2 and H+ Concentration on Alveolar Ventilation)
図 42-3 (Figure 42-3) では、血液中のPco2 および血液 pH(H+ 濃度の逆対数的指標)が肺換気に及ぼす影響を定量的に示しています。特に、正常範囲の35〜75 mm HgのPco2 の増加が肺換気を大きく増加させる様子が示されています。これは、CO2 の変化が呼吸の制御に与える巨大な影響を示しています。一方で、正常な血液pH範囲(7.3〜7.5)での呼吸の変化は、CO2 に比べて10%以下の影響しかありません。
酸素の直接的影響 (Direct Effect of Oxygen)
酸素の濃度変化は、呼吸中枢自体にはほとんど直接的な影響を与えません。酸素の変化は主に末梢化学受容体 (peripheral chemoreceptors) に作用し、これらの受容体が適切な神経信号を呼吸中枢に送って呼吸を制御します。
ヘモグロビン-酸素バッファーシステム (Hemoglobin-Oxygen Buffer System) は、肺のPo2 が60 mm Hg から1000 mm Hg に変化しても、ほぼ正常な量の酸素を組織に供給します。したがって、酸素の供給は、肺換気が正常の半分以下から20倍以上に変化しても適切に行われることができます。しかし、CO2 は異なり、血液および組織のPco2 は肺換気の率に逆相関するため、進化の過程でCO2 が呼吸の主要な制御因子となっています。
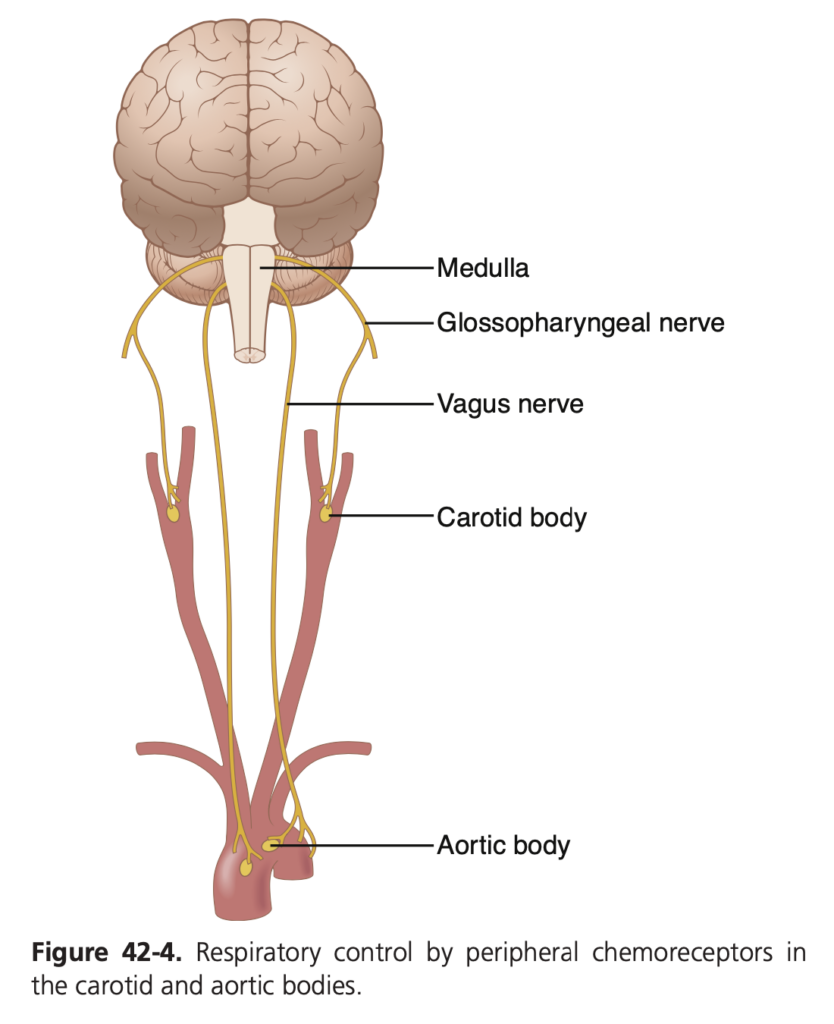
末梢化学受容体系 (Peripheral Chemoreceptor System) – 酸素の呼吸制御における役割 (Role of Oxygen in Respiratory Control)
呼吸中枢による制御に加え、末梢化学受容体系も呼吸を制御するための別のメカニズムを提供します。末梢化学受容体 (chemoreceptors) は、脳の外部にあるいくつかの領域に位置しており、特に血液中の酸素 (O2) の変化を検出するのに重要です。これらの受容体は、CO2 やH+ 濃度の変化にもある程度反応しますが、主に酸素濃度に反応します。
- 内頸動脈小体 (Carotid Bodies): 内頸動脈の分岐部に位置し、ヘーリング神経 (Hering’s nerves) を通じて咽頭舌神経 (glossopharyngeal nerves) に接続され、延髄の背側呼吸領域 (dorsal respiratory area) に信号を送ります。
- 大動脈小体 (Aortic Bodies): 大動脈の弓部に位置し、迷走神経 (vagus nerves) を通じて延髄の背側呼吸領域に信号を送ります。
これらの化学受容体は、血液の流れが非常に速く、ほぼ動脈血に常にさらされているため、酸素の供給が最適化されています。
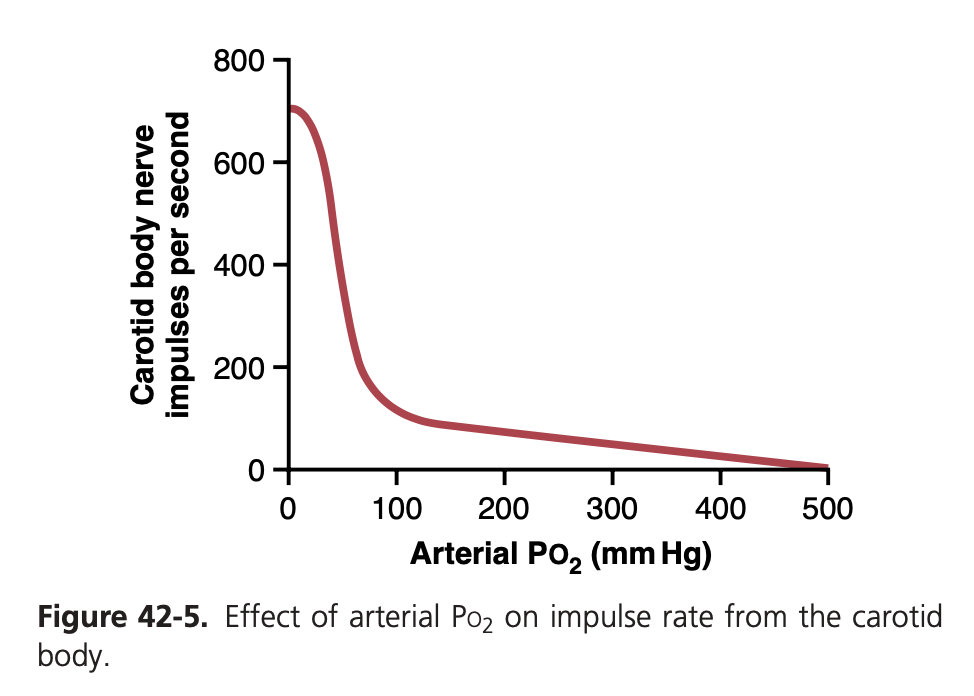
酸素不足による化学受容体の刺激 (Decreased Arterial Oxygen Stimulates the Chemoreceptors)
血液中の酸素濃度が正常値を下回ると、化学受容体は強く刺激されます。酸素濃度が60 mm Hg から30 mm Hg の範囲で特に敏感に反応し、ヘモグロビンの酸素飽和度が急激に低下します。
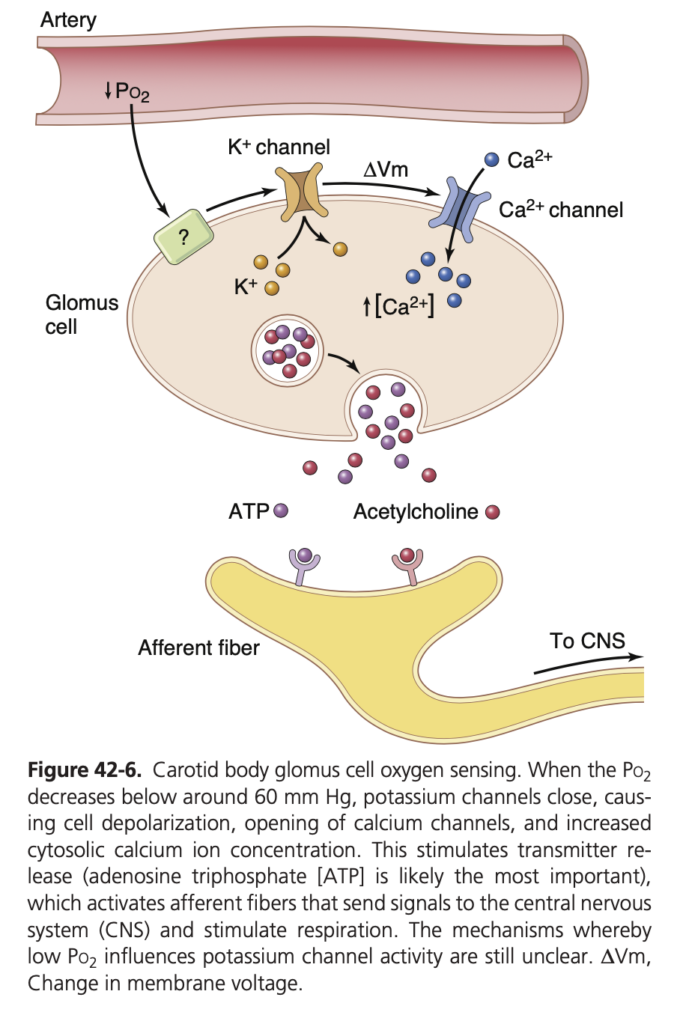
酸素欠乏による化学受容体の基本メカニズム (Basic Mechanism of Stimulation of the Chemoreceptors by O2 Deficiency)
酸素欠乏が内頸動脈小体や大動脈小体の神経末端をどのように刺激するかは完全には理解されていませんが、これらの小体には「グロムス細胞 (glomus cells)」と呼ばれる特徴的な細胞があり、神経末端と直接または間接的にシナプスを形成します。グロムス細胞は酸素感受性のカリウムチャネルを持ち、酸素濃度が低下するとカリウムチャネルが不活化し、細胞膜が脱分極します。この過程でカルシウムチャネルが開き、細胞内カルシウム濃度が増加します。増加したカルシウムイオンが神経伝達物質の放出を促し、感覚神経が中枢神経系に信号を送って呼吸を刺激します。
CO2 と H+ の化学受容体への刺激 (Increased CO2 and H+ Concentration Stimulates the Chemoreceptors)
CO2 やH+ の濃度が増加すると、化学受容体も刺激され、間接的に呼吸活動が増加します。しかし、CO2 やH+ の直接的な影響は、化学受容体を介した影響よりも約7倍強力です。また、末梢化学受容体による刺激は中枢刺激の5倍早く発生するため、運動の開始時におけるCO2 に対する応答の迅速さを高めるために重要です。
低動脈Po2が肺換気に及ぼす影響 (Effect of Low Arterial Po2 on Alveolar Ventilation)
図 42-7 (Figure 42-7) では、動脈中のPco2 とH+ 濃度が正常レベルに保たれている場合の低動脈Po2 が肺換気に与える影響が示されています。つまり、この図では、低酸素によって化学受容体が引き起こす換気ドライブのみが活性化されています。図は、動脈Po2 が100 mm Hg より高い場合、換気にほとんど影響がないことを示しています。しかし、Po2 が100 mm Hg 以下になると、Po2 が60 mm Hg に低下した際に換気が約2倍になり、非常に低いPo2 値では換気が最大5倍に増加する可能性があります。この条件下では、低動脈Po2 が換気プロセスを強力に駆動することが明らかです。
動脈Po2 が60〜80 mm Hg 以上である場合の低酸素の影響が比較的軽微であるため、健康な人々の海面レベルでの換気調節は主にPco2 およびH+ の反応によって行われます。
低酸素の慢性的な呼吸が呼吸をさらに刺激する—「順応現象」 (Chronic Breathing of Low O2 Stimulates Respiration Even More—The Phenomenon of “Acclimatization”)
登山者は、急激に山を登るのではなく、数日かけて徐々に登ると、はるかに深い呼吸をするようになり、低い大気中の酸素濃度に耐えられることを発見しています。この現象は「順応 (acclimatization)」と呼ばれます。
順応の理由は、2〜3日以内に脳幹の呼吸中枢がPco2 およびH+ の変化に対する感受性を約80%失うためです。したがって、通常は呼吸の増加を抑制するはずのCO2 の過剰な排出が行われず、低酸素が呼吸系をより高いレベルの肺換気に駆動します。急性の低酸素暴露後の70% の換気増加に対し、2〜3日後には肺換気が400%〜500% 増加し、登山者に追加の酸素を供給するのに大いに役立ちます。
Pco2、pH、Po2 の複合的な影響 (Composite Effects of Pco2, pH, and Po2 on Alveolar Ventilation)
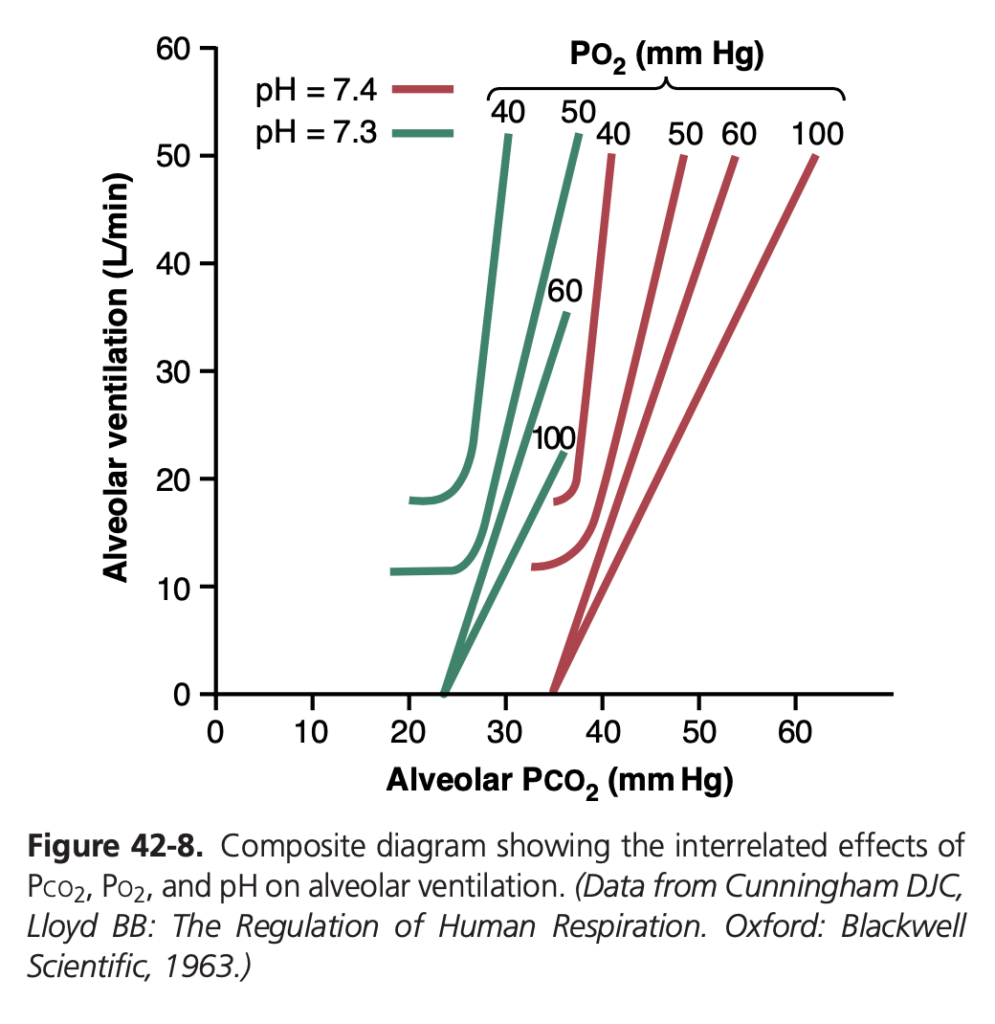
図 42-8 (Figure 42-8) では、Po2、Pco2、およびpH がどのように肺換気に複合的に影響を与えるかが概要として示されています。この図を理解するためには、まず4つの赤い曲線を観察します。これらの曲線は動脈Po2 の異なるレベル(40、50、60、100 mm Hg)で記録されており、各曲線でPco2 が低いレベルから高いレベルに変化しています。したがって、これらの赤い曲線群は、肺換気に対するPco2 とPo2 の複合的な影響を表しています。
次に緑の曲線に注目します。赤い曲線が血液pH 7.4 で測定されたのに対し、緑の曲線はpH 7.3 で測定されました。これにより、2つの異なるpH 値でのPco2 とPo2 の肺換気に対する複合的な影響が示されます。さらに、高pH では曲線が右に、低pH では左に移動します。この図を使用することで、ほとんどのPco2、Po2、および動脈pH の組み合わせに対する肺換気のレベルを予測することができます。
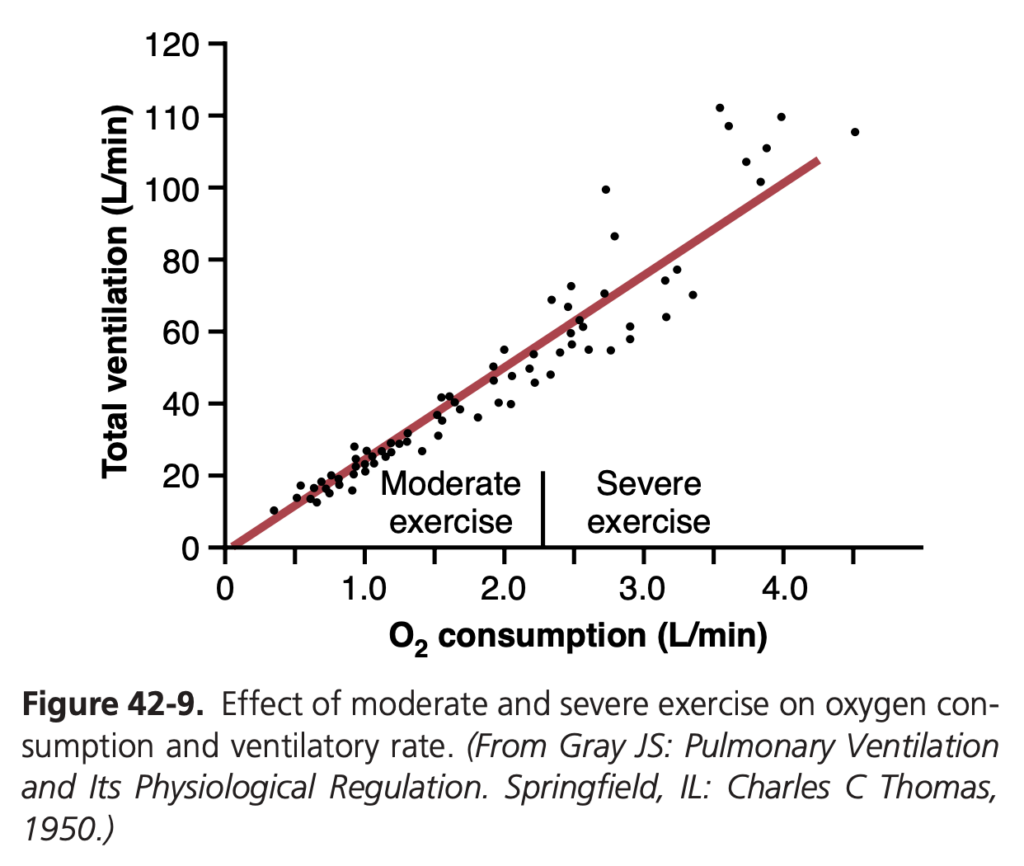
運動中の呼吸調節 (Regulation of Respiration During Exercise)
運動中には、酸素消費と二酸化炭素の生成が最大で20倍まで増加します。それにもかかわらず、健康なアスリートでは、図 42-9 に示されているように、肺換気は通常、酸素代謝の増加とほぼ正確に一致して増加します。動脈のPo2、Pco2、pHはほぼ正常のままです。
運動中の換気増加の原因
運動中の換気増加は、血中のCO2とH+の増加やO2の減少に起因すると思われがちですが、動脈のPco2、pH、Po2 の測定値は運動中に有意に変化しないため、これらの変化が運動中の強い換気を直接引き起こすことはありません。運動中の強い換気の主な原因の一つは、運動を始める際に脳が運動筋に信号を送ると同時に、脳幹にある呼吸中枢にも副次的な信号を送ることによって呼吸中枢が興奮することです。この作用は、運動中に血圧が同時に上昇するのと類似しています。
実際、運動を始めると、血液化学物質が変化する前に、換気の増加の大部分が即座に始まります。これは、呼吸中枢が筋肉の収縮を引き起こす信号と同時に直接神経信号を送ることによって、ほとんどの換気増加がもたらされると考えられます。
運動中の化学的および神経的要因の相互関係
運動中には、直接的な神経信号が呼吸中枢をほぼ適切な量で刺激し、運動に必要な追加の酸素供給と二酸化炭素の排出を実現します。しかし、神経的な呼吸調節信号が強すぎたり弱すぎたりする場合、化学的要因が体液のO2、CO2、およびH+ 濃度を可能な限り正常に保つために最終的な調整を行います。
図 42-10 では、このプロセスが示されています。下の曲線は運動1分間の肺換気の変化を示し、上の曲線は動脈Pco2の変化を示しています。運動の開始時には、動脈Pco2 の増加がないまま肺換気がほぼ即座に増加し、換気が通常大きすぎるため、初期には動脈Pco2 が正常以下に減少することが多いです。このように、換気が血液中のCO2 の増加よりも先行して増加する理由は、脳が運動の開始時に「予測的」な呼吸刺激を提供し、必要以上に換気が行われるからです。しかし、運動開始から30〜40秒後には、活動する筋肉から血液に放出されるCO2 の量が増加した換気速度にほぼ一致し、動脈Pco2 はほぼ正常に戻ります。
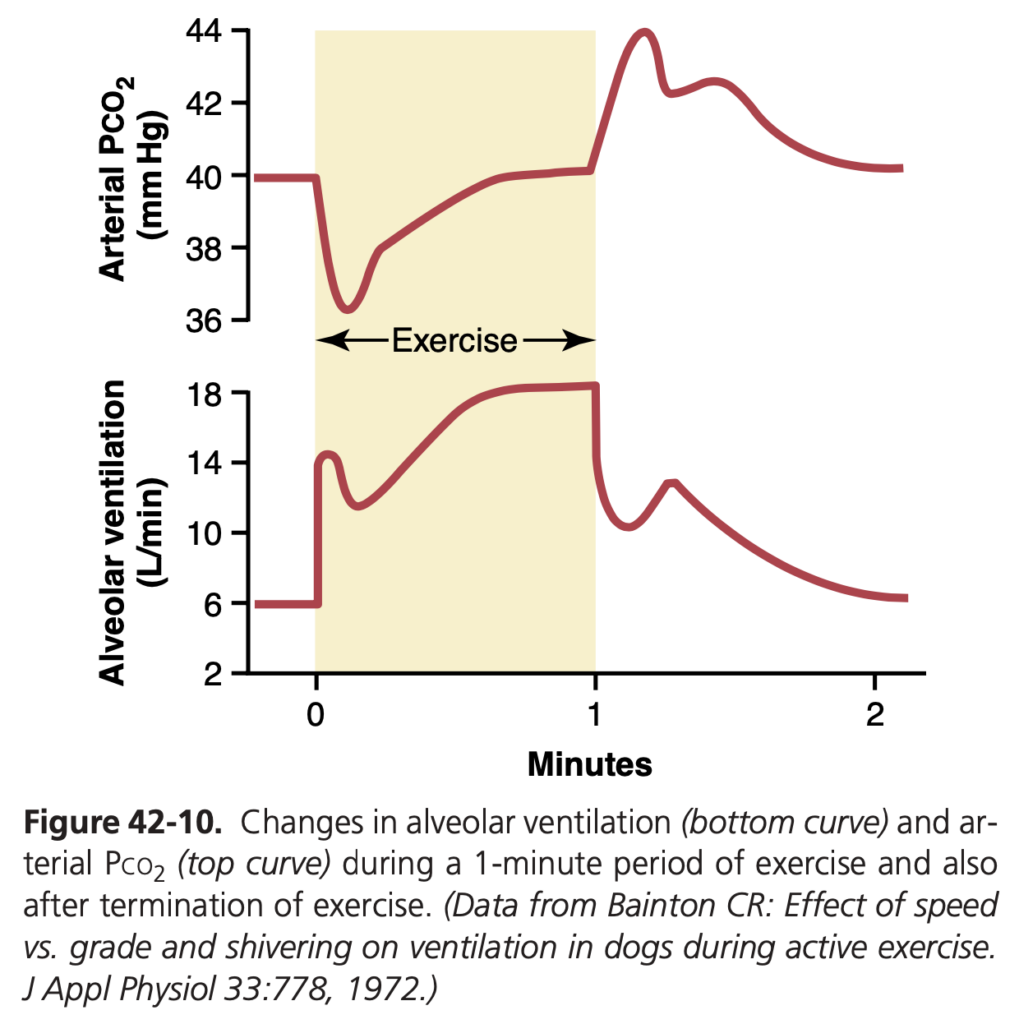
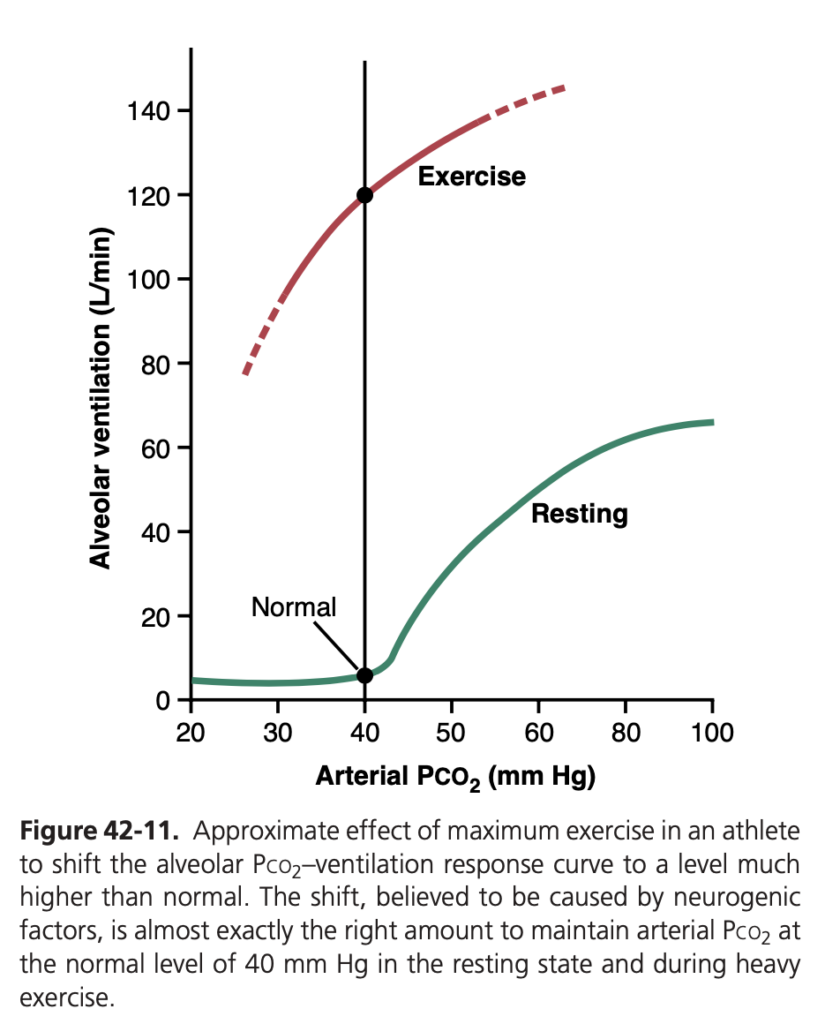
図 42-11 では、運動中の呼吸調節が別の方法で要約されています。下の曲線は、安静時における異なる動脈Pco2 のレベルが肺換気に与える影響を示し、上の曲線は重い運動中の神経的ドライブによる換気曲線のシフトを示しています。両曲線の点は、安静時と運動時の動脈Pco2 を示しており、どちらのケースでもPco2 は正常レベルの40 mm Hg にあります。神経的要因は曲線を約20倍上にシフトさせ、換気はCO2 の排出速度にほぼ一致し、動脈Pco2 を正常値に保ちます。また、運動中に動脈Pco2 が40 mm Hg から変化する場合、Pco2 が40 mm Hg より高いと換気を刺激し、40 mm Hg より低いと抑制する追加的な効果があります。
運動中の換気の神経的制御は学習された反応の一部かもしれない
多くの実験は、図 42-11 に示されたように、運動中の換気反応曲線をシフトさせる脳の能力が、少なくとも一部は学習された反応であることを示唆しています。つまり、運動の繰り返しによって、脳は血液Pco2 を正常レベルに保つために必要な適切な信号を提供する能力が次第に向上します。さらに、脳皮質もこの学習に関与している可能性があり、脳皮質をブロックする実験では学習された反応がブロックされることが示されています。
呼吸に影響を与えるその他の要因 (Other Factors That Affect Respiration)
気道における刺激受容体の影響
気管、気管支、細気管支の上皮には、肺刺激受容体 (pulmonary irritant receptors) と呼ばれる感覚神経末端が分布しています。これらの受容体は多くの因子によって刺激され、咳嗽やくしゃみを引き起こすほか、喘息や慢性閉塞性肺疾患 (COPD) などの疾患を持つ人では気管支収縮を引き起こすことがあります。
肺J受容体の機能
肺の毛細血管と接触している肺胞壁には、J受容体 (J receptors) と呼ばれる感覚神経末端が存在します。これらの受容体は、肺毛細血管が血液で充満する場合や、肺水腫が発生する場合に特に刺激されます。J受容体の機能的役割は明確ではありませんが、その刺激は呼吸困難感を引き起こす可能性があります。
脳浮腫による呼吸中枢の抑制
脳震盪などによって急性の脳浮腫が発生すると、呼吸中枢の活動が抑制されることがあります。例えば、頭部が固い物体にぶつかると、損傷した脳組織が腫れ、脳の血管が圧迫されることで脳血流が部分的に遮断されることがあります。
脳浮腫による呼吸抑制は、濃度の高いマニトール溶液などの高浸透圧液体を静脈注射することで一時的に改善されることがあります。これにより、脳内の液体が浸透圧的に除去され、頭蓋内圧が軽減されることで、数分以内に呼吸が再開することがあります。
麻酔薬や麻薬の過剰摂取
麻酔薬や麻薬の過剰摂取は、呼吸抑制や呼吸停止の最も一般的な原因の一つです。例えば、ナトリウムペントバルビタール (sodium pentobarbital) は、多くの他の麻酔薬よりも呼吸中枢を著しく抑制します。一時期、モルヒネは麻酔薬として使用されていましたが、現在は麻酔の補助剤としてのみ使用されており、脳皮質の麻酔作用は少ないものの、呼吸中枢を強く抑制します。
麻薬による呼吸抑制のため、世界中で高い割合の致命的な薬物過剰摂取が報告されています。アメリカ合衆国では、2017年に約70,000人が薬物過剰摂取によって死亡しました。
周期的呼吸
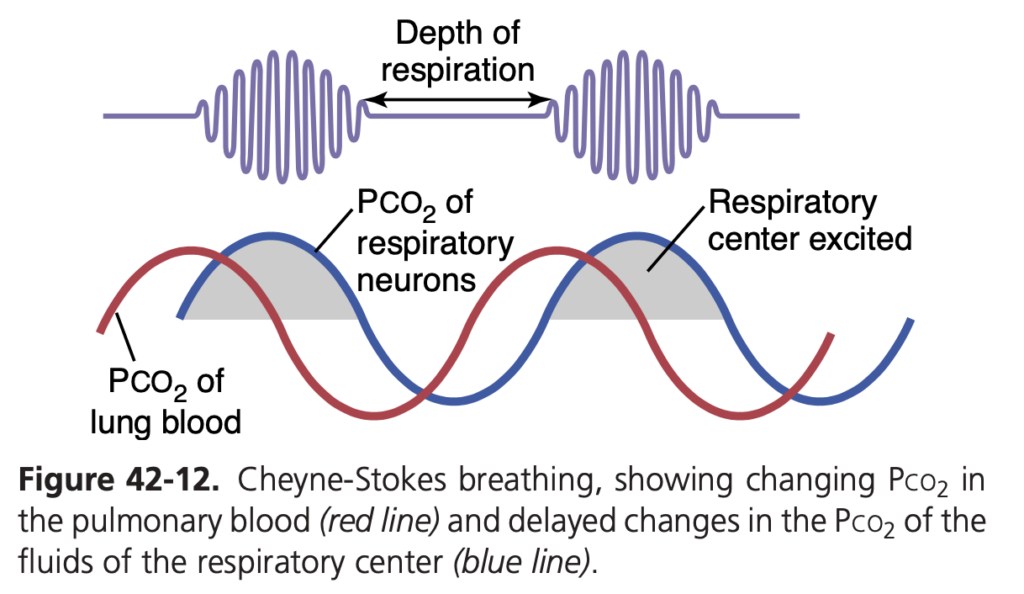
周期的呼吸 (periodic breathing) という異常な呼吸パターンは、いくつかの疾患で見られます。周期的呼吸では、短時間の深い呼吸の後に、ほとんど呼吸がなくなるまたはわずかに呼吸する期間が続き、このサイクルが繰り返されます。チェーン・ストークス呼吸 (Cheyne-Stokes breathing) は、この周期的呼吸の一例で、約40〜60秒ごとに呼吸が徐々に増加し、その後減少するパターンが見られます(図 42-12 参照)。
チェーン・ストークス呼吸の基本的メカニズム
チェーン・ストークス呼吸の基本的な原因は次の通りです。過剰に呼吸すると、肺から血液中のCO2が過剰に排出され、同時に血液中のO2が増加します。この変化した肺血液が脳に送られ、過剰な換気を抑制するまでに数秒かかります。この間に、過剰な換気がさらに続きます。その結果、過剰に換気された血液が脳に到達すると、呼吸中枢が過剰に抑制され、逆のサイクルが始まります。すなわち、CO2 が増加し、O2 が減少します。この変化にも脳が応答するのに数秒かかり、その後、再び呼吸が増加し、このサイクルが繰り返されます。
その他の呼吸に影響を与える要因
チェーン・ストークス呼吸の基本的な原因
チェーン・ストークス呼吸(Cheyne-Stokes breathing)は、すべての人において基本的に発生しますが、正常な状態ではこのメカニズムは強く抑制されています。つまり、血液と呼吸中枢の制御領域には、大量の溶解CO2および化学的に結合したO2が存在するため、通常、肺ではCO2が過剰に蓄積されたり、O2が十分に低下することがなく、周期的呼吸の次のサイクルを引き起こすことはありません。しかし、以下の2つの条件では抑制因子が打破され、チェーン・ストークス呼吸が発生します。
- 血液の肺から脳への輸送遅延: 血液が肺から脳に輸送されるのに時間がかかる場合、肺胞内のCO2およびO2の変化が通常よりも長く続くことがあります。この場合、肺胞および肺血液のガス貯蔵能力を超えて、さらに数秒後に周期的呼吸の駆動が強くなり、チェーン・ストークス呼吸が始まります。このタイプのチェーン・ストークス呼吸は、血流が遅いために肺から脳への血液ガス輸送が遅れる重度の心不全患者によく見られます。慢性心不全患者では、チェーン・ストークス呼吸が数ヶ月にわたって発生することがあります。
- 呼吸制御領域の負のフィードバックゲインの増加: 呼吸制御領域での負のフィードバックゲインが増加すると、血液中のCO2またはO2の変化が通常よりも大きな変化を引き起こします。例えば、Pco2が3 mm Hg上昇したときに、通常は換気が2〜3倍増加しますが、同じ3 mm Hgの上昇が10〜20倍の換気増加を引き起こすことがあります。このタイプのチェーン・ストークス呼吸は、脳の呼吸中枢がダメージを受けた場合に見られます。脳の損傷が呼吸駆動を完全に停止させることがあり、その後、血中CO2の急激な増加が強力な呼吸駆動を引き起こします。このタイプのチェーン・ストークス呼吸は、脳機能不全による死の前兆であることが多いです。チェーン・ストークス呼吸中の肺および呼吸中枢Pco2の変化を示した典型的な記録は図 42-12 に示されています。肺血液のPco2は呼吸ニューロンのPco2に先行して変化し、呼吸の深さは肺血液のPco2ではなく、脳内のPco2に対応しています。
睡眠時無呼吸症候群 (Sleep Apnea)
睡眠時無呼吸症候群 (sleep apnea) とは、自然呼吸が停止する状態を指します。正常な睡眠中にも時折無呼吸が見られますが、睡眠時無呼吸症候群のある人々では、無呼吸の頻度と持続時間が大幅に増加し、10秒以上の無呼吸が1晩に300〜500回発生することがあります。睡眠時無呼吸症候群の原因には、上気道(特に咽頭)の閉塞または中枢神経系の呼吸駆動の障害が含まれます。
- 閉塞性睡眠時無呼吸 (Obstructive Sleep Apnea): 咽頭の筋肉が通常、呼吸のために気道を開けておく役割を果たしていますが、睡眠中にこれらの筋肉が弛緩すると、一部の人々では咽頭が完全に閉じ、空気が肺に流れなくなります。閉塞性睡眠時無呼吸では、大きな音でいびきをかいたり、呼吸が困難になったりします。いびきが続いた後、長い無呼吸の静かな期間があり、この無呼吸によりPO2が大幅に低下し、Pco2が増加します。これにより呼吸が刺激され、突然の呼吸努力がいびきや喘ぎを引き起こします。このような無呼吸と呼吸困難が夜間に何百回も繰り返され、睡眠が断続的で落ち着かないものになります。その結果、昼間の過度の眠気、交感神経活動の増加、高い心拍数、肺および全身の高血圧、心血管疾患のリスクが大幅に増加します。閉塞性睡眠時無呼吸は、通常、肥満の高齢者に見られ、咽頭の軟部組織に脂肪が蓄積して気道が狭くなるか、首の過剰な脂肪が咽頭を圧迫することによって発生します。まれに、鼻の閉塞、大きな舌、肥大した扁桃腺、または上顎の特定の形状が原因で、吸気時の空気流の抵抗が大幅に増加することがあります。閉塞性睡眠時無呼吸の治療法としては、(1)喉の後部の脂肪組織を除去する手術(口蓋垂咽頭形成術(uvulopalatopharyngoplasty))、肥大した扁桃腺やアデノイドの除去、気道をバイパスするための気管切開術(tracheostomy)など、(2)持続的陽圧呼吸(CPAP)による鼻への換気が含まれます。
- 中枢性睡眠時無呼吸 (Central Sleep Apnea): 中枢性睡眠時無呼吸では、中枢神経系からの呼吸筋への駆動が一時的に消失します。中枢性睡眠時無呼吸の原因には、呼吸中枢の損傷や呼吸神経筋装置の異常が含まれます。中枢性睡眠時無呼吸の患者は、覚醒時でも換気が減少することがありますが、通常の自発的呼吸は可能です。睡眠中には呼吸障害が悪化し、無呼吸の頻度が増し、PO2が低下し、Pco2が増加します。これにより呼吸が再び刺激されますが、結果として不安定な呼吸が生じ、閉塞性睡眠時無呼吸に類似した臨床的特徴が見られます。中枢性睡眠時無呼吸の原因は多くの場合不明ですが、脳卒中やその他の疾患が呼吸中枢のCO2やH+に対する反応性を低下させることがあります。この疾患の患者は、少量の鎮静剤や麻薬に対して非常に敏感で、呼吸中枢のCO2に対する反応性がさらに低下します。呼吸中枢を刺激する薬物が有用な場合もありますが、通常はCPAPによる換気が必要です。一部の症例では、閉塞性と中枢性のメカニズムが組み合わさっている「混合型」睡眠時無呼吸が見られることがあります。この「混合型」睡眠時無呼吸は、睡眠時無呼吸症候群の約15%を占めており、純粋な「中枢性」睡眠時無呼吸は1%未満です。最も一般的な原因は、上気道の閉塞です。
自発的呼吸の制御
これまで、主に呼吸の無意識的な制御システムについて説明してきましたが、呼吸は短期間で意図的に制御することができます。例えば、呼吸を抑制することによって、Pco2、pH、Po2の血中濃度に重大な変化を引き起こすことがあります。実際、静止状態での自主的な息止め(アプニア)の世界記録は11分54秒です。純酸素で過換気した後の息止めでは、24分以上の息止めが可能になります。超エリートアプニア競技者は、呼吸衝動を抑制することによって、酸素飽和度が約50%まで低下することができ、意識喪失が息止めの持続時間を制限します。

コメント